
Journal of Tropical Oceanography >
Adaptive mechanisms of iron limitation on the marine Synechococcus based on comparative genomics
Copy editor: LIN Qiang
Received date: 2023-03-23
Revised date: 2023-04-28
Online published: 2023-05-08
Supported by
National Natural Science Foundation of China(41890801)
International Cooperation in Science and Technology Innovation between Governments(2022YFE0136500)
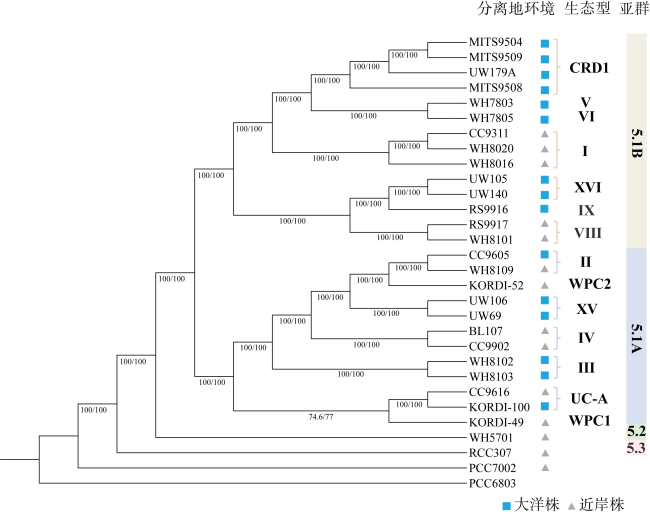
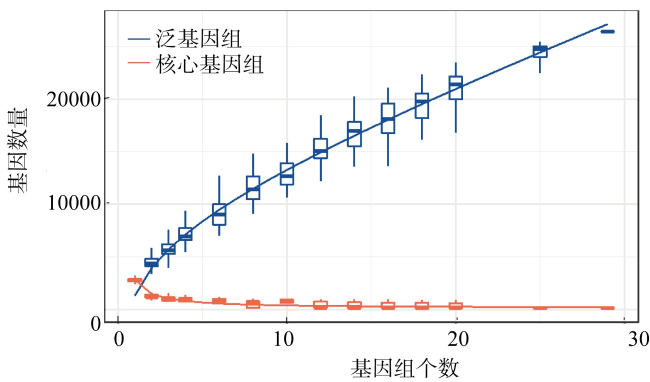
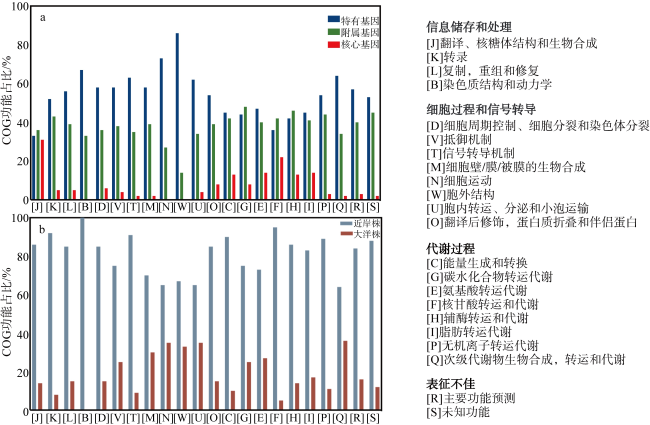
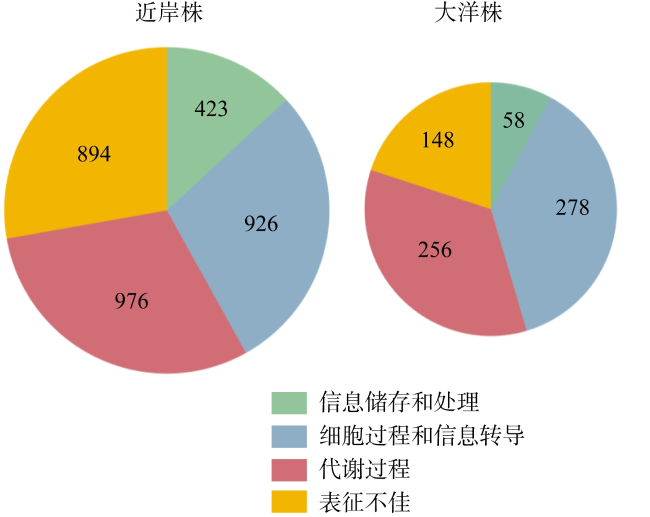
Low solubility and limited source of iron in seawater are the main growth limiting factors for phytoplankton in 40%of the global ocean. Marine Synechococcus is a picocyanobacteria with global distribution and is one of the most important contributors to marine prime productivity. Affected by the source and concentration of iron in coastal and oceanic environments, marine Synechococcus has different adaptive mechanisms to iron limitation. In this study, we performed a comparative genomic analysis of 29 whole genome sequenced marine Synechococcus. The results showed that marine Synechococcus had high genetic diversity and belonged to four genera of Cyanobiaceae under the GTDB (Genome Taxonomy Database) taxonomy, and these four genera corresponded well to the subtype under NCBI (National Center for Biotechnology Information) taxonomy. The results of functional annotation showed that the number and types of unique genes differed between coastal and oceanic Synechococcus, and unique gene of coastal strains are more involved in inorganic ion transport and metabolism. Further analysis of iron limitation-related genes in marine Synechococcus revealed that the coastal strain had stronger abilities of iron uptake, homeostasis regulation and storage than the oceanic strains, and had a better ability to sense and respond to environmental changes. In this study, we reviewed the evolutionary relationships and taxonomic positions of marine Synechococcus using comparative genomics, and identified the differences in the genome and adaptive mechanisms of coastal and oceanic strains in response to iron limitation, to provide a basis for better understanding of the environmental adaptation of marine Synechococcus.
MU Rong , ZHU Zhu , ZHANG Ruifeng . Adaptive mechanisms of iron limitation on the marine Synechococcus based on comparative genomics[J]. Journal of Tropical Oceanography, 2023 , 42(6) : 89 -100 . DOI: 10.11978/2023036
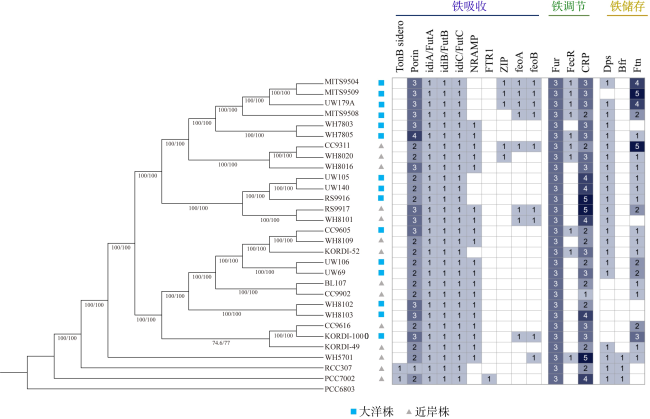
表1 本研究中铁限制相关功能基因Tab. 1 Functional genes related to iron limitation |
| 基因分类 | 基因 | 基因描述 | KEGG 编号 | |
|---|---|---|---|---|
| 铁吸收 | 铁载体相关基因 | TonB sidero | TonB dependent receptor | K02014 |
| 孔蛋白基因 | Porin | Outer membrane protein insertion porin family | K07277 | |
| Fe3+转运基因 | idiA/FutA | Iron deficiency induced protein A / iron(Ⅲ) transport system substrate-binding protein | K02012 | |
| idiB/FutB | ABC-type Fe3+ transport system permease component / iron(Ⅲ) transport system permease protein | K02011 | ||
| idiC/FutC | ATP binding component / iron(Ⅲ) transport system ATP-binding protein | K02010 | ||
| 二价金属转运基因 | NRAMP | Natural resistance associated macrophage protein, Nramp | K03322 | |
| FTR1 | FTR1 family iron permease | K07243 | ||
| ZIP | ZRT / IRT-like Protein | K14709 | ||
| Fe2+转运基因 | FeoA | Ferrous iron transport protein A | K04758 | |
| FeoB | Ferrous iron transport protein B | K04759 | ||
| 铁稳态 | — | Fur | Ferric uptake regulator | K03711 |
| FecR | Periplasmic ferric-dicitrate binding protein FerR | K07165 | ||
| CRP | Cyclic AMP receptor protein | K10914 | ||
| 铁储存 | 铁储存蛋白 | Dps | DNA-binding stress protein | K04047 |
| Bfr | Bacterioferritin | K03594 | ||
| Ftn | Ferritins | K02217 | ||
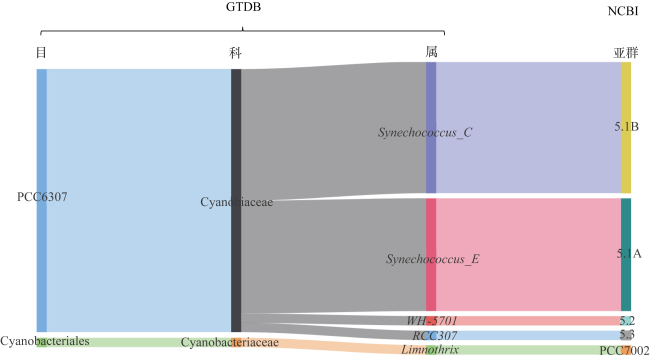
图2 海洋聚球藻GTDB分类法与NCBI分类法的对应关系用GTDB-Tk对来自3个亚群的29株海洋聚球藻基因组进行物种注释, 确定其在GTDB分类法(Release 07-RS207)中所属的目(order)、科(family)和属(genus)的分类地位, 通过桑基图来表示海洋聚球藻GTDB分类法和NCBI分类法中亚群之间的对应关系 Fig. 2 Relationship between the GTDB taxonomy and the NCBI taxonomy of marine Synechococcus |
表2 本研究中29株海洋聚球藻的基因组信息Tab. 2 Genome features of the 29 analyzed marine Synechococcus strains |
| 分离地点 | 藻株 | 基因组编号 | 类型 | 基因组大小/bp | GC含量/% | 完整度/% | 污染度/% | |
|---|---|---|---|---|---|---|---|---|
| 大西洋马尾藻海 | WH8102 | GCA_000195975.1 | 大洋 | 2434428 | 59.4 | 99.46 | 0 | |
| WH7803 | GCA_000063505.1 | 大洋 | 2366980 | 60.2 | 99.18 | 0 | ||
| WH8103 | GCA_001182765.1 | 大洋 | 2429688 | 59.5 | 99.73 | 0.27 | ||
| UW69 | GCA_900474185.1 | 大洋 | 2372121 | 58.7 | 100 | 0 | ||
| UW140 | GCA_900474295.1 | 大洋 | 2704142 | 57.8 | 99.73 | 0 | ||
| UW105 | GCA_900473935.1 | 大洋 | 2659417 | 57.5 | 99.46 | 0 | ||
| UW106 | GCA_900474015.1 | 大洋 | 2479535 | 58 | 99.73 | 0.27 | ||
| WH8109 | GCA_000161795.2 | 大洋 | 2111515 | 60.1 | 99.46 | 0 | ||
| 赤道太平洋 | UW179A | GCA_900473965.1 | 大洋 | 3057835 | 54.5 | 100 | 0.41 | |
| MITS9508 | GCA_001632165.1 | 大洋 | 2502434 | 56.1 | 100 | 0.27 | ||
| MITS9509 | GCA_001631935.1 | 大洋 | 3087928 | 55.4 | 99.18 | 0.82 | ||
| MITS9504 | GCA_001632105.1 | 大洋 | 3087293 | 55.4 | 99.73 | 1.09 | ||
| 东太平洋加利福尼亚海流 | CC9605 | GCA_000012625.1 | 近岸 | 2510659 | 59.2 | 99.73 | 0 | |
| CC9311 | GCA_000014585.1 | 近岸 | 2606748 | 52.5 | 99.73 | 0 | ||
| CC9902 | GCA_000012505.1 | 近岸 | 2234828 | 54.2 | 99.46 | 0 | ||
| 中国东海 | KORDI-52 | GCA_000737595.1 | 近岸 | 2572069 | 59.1 | 100 | 0 | |
| KORDI-49 | GCA_000737575.1 | 近岸 | 2585813 | 61.4 | 99.37 | 0 | ||
| 地中海 | RCC307 | GCA_000063525.1 | 近岸 | 2224914 | 60.8 | 99.64 | 0 | |
| BL107 | GCA_000153805.1 | 近岸 | 2285034 | 54.2 | 99.46 | 0 | ||
| 红海亚喀巴湾 | RS9916 | GCA_000153825.1 | 近岸 | 2664873 | 59.8 | 99.73 | 0 | |
| RS9917 | GCA_000153065.1 | 近岸 | 2584918 | 64.5 | 99.46 | 0 | ||
| 波多黎马格耶斯岛 | PCC7002 | GCA_000019485 | 近岸 | 3409935 | 49.2 | 100 | 0 | |
| 西太平洋 | KORDI-100 | GCA_000737535.1 | 大洋 | 2789000 | 57.5 | 99.46 | 0 | |
| 新英格兰大陆架 | WH8020 | GCA_001040845.1 | 近岸 | 2661166 | 53.1 | 99.73 | 0.27 | |
| 伍兹霍尔沿海海水 | WH8101 | GCA_004209775.1 | 近岸 | 2630292 | 63.3 | 100 | 0.82 | |
| 北大西洋潮间带 | WH8016 | GCA_000230675.2 | 近岸 | 2694843 | 54.1 | 99.18 | 0 | |
| 北大西洋 | WH7805 | GCA_000153285.1 | 大洋 | 2627046 | 57.6 | 99.73 | 0 | |
| 北大西洋长岛海峡 | WH5701 | GCA_000153045 | 近岸 | 3280236 | 65.4 | 99.18 | 7.84 | |
| 东太平洋 | CC9616 | GCA_000515235.1 | 大洋 | 2645910 | 56.6 | 99.46 | 0.27 | |
表3 近岸和大洋聚球藻对铁限制的适应机制差异Tab. 3 Differences in adaptive mechanisms for iron limitation of coastal and oceanic Synechococcus |
| 相关基因/蛋白 | 海洋聚球藻 | ||
|---|---|---|---|
| 近岸株 | 大洋株 | ||
| 铁吸收 | 铁载体 | 合成和利用铁载体的能力较强 | 合成和利用铁载体的能力较弱 |
| 孔蛋白 | 存在 | 存在 | |
| Fe(Ⅲ)转运蛋白 | 存在, 基因表达量相对较低 | 存在, 基因表达量相对较高 | |
| Fe(Ⅱ)特异性转运蛋白 | 普遍存在 | 除生态型CRD1中不存在 | |
| 铁稳态 | Fur家族(FurA、Zur、PerR) | 存在, 具有更复杂的调节能力 | 存在 |
| FecR | 存在 | 仅在生态型CRD1中广泛分布 | |
| CRP | 存在 | 存在 | |
| 铁储存 | Ftn | 拷贝数较多 | 仅在生态型CRD1中拥有多拷贝数 |
| Bfr | 存在 | 不存在 | |
| Dps | 存在 | 存在 | |
| [1] |
彭铭烨, 黄婷, 张晓娟, 等, 2023. 比较基因组学解析谷物醋醋醅中巴氏醋杆菌和欧洲驹形杆菌的功能差异[J]. 微生物学报, 63(2): 638-655.
|
| [2] |
郑强, 贺博闻, 史文卿, 等, 2023. 海洋超微型蓝细菌聚球藻的生态学研究进展[J]. 厦门大学学报(自然科学版), 62(3): 301-313.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}