
Journal of Tropical Oceanography >
Pattern recognition of Sthenoteuthis oualaniensis based on BPNN about momentum and self-adaption
Copy editor: LIN Qiang
Received date: 2020-11-17
Revised date: 2021-02-27
Online published: 2021-03-04
Supported by
National Key R&D Program of China(2019YFD0901404)
National Nature Science Foundation of China(41876141)
The Program for Professor of Special Appointment (Eastern Scholar) at Shanghai Institutions of Higher Learning under contract(0810000243)
Capacity building project of local universities of Shanghai Municipal Science and Technology Committee(20050501800)
Shanghai Science and technology innovation action plan(19DZ1207502)
Copyright
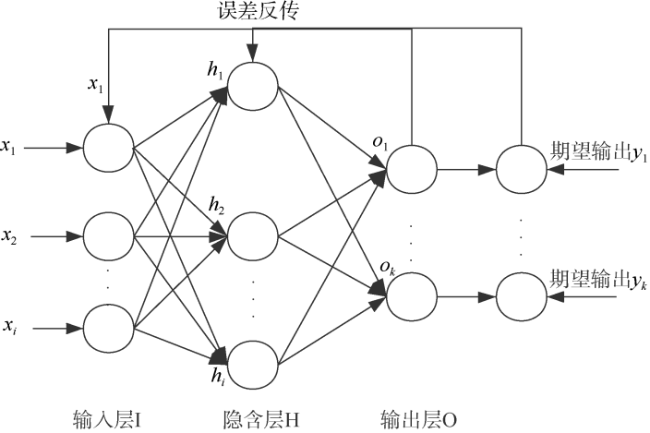
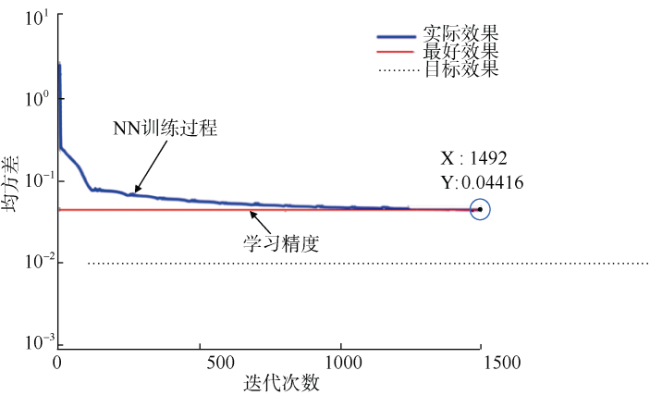
In recent years, computer pattern recognition technology has been used in biometric identification for its accurate and rapid recognition capability. In this paper, pattern recognition of beak and mantle length of Sthenoteuthis oualaniensis in the northwestern Indian Ocean, Middle East Pacific and South China Sea was carried out by using momentum adaptive back propagation (BP) neural networks (BPNN) based on MATLAB software. The results showed that the convergence error of the trained neural network was only 4.416×10-2, and the recognition rate of beak of S. oualaniensis was significantly improved by adding momentum and adaptive learning rate to the BPNN. The correct recognition rates were 100% in the northwestern Indian Ocean, 88.89% in the Middle East Pacific and 94.12% in the South China Sea, with a total success rate of 93.24%, which indicates that the external morphology of beak of S. oualaniensis and mantle length can be used to distinguish different geographical populations. The total recognition rates of other BPNN learning algorithms of gradient descent, single momentum and single adaptive method were 74.32%, 77.03% and 87.84%, respectively. The recognition effect of this study was stable, and the recognition rate of large sample training set was as high as 92.77%, which provides a new method for the identification of cephalopod population.
YANG Liuqinqing , CHU Moxian , LIU Bilin , KONG Xianghong . Pattern recognition of Sthenoteuthis oualaniensis based on BPNN about momentum and self-adaption[J]. Journal of Tropical Oceanography, 2021 , 40(6) : 102 -110 . DOI: 10.11978/2020135
表1 标准化后的极值和所属海域Tab. 1 The extreme value and its sea area after standardization |
| 形态参数 | 极大值 | 极小值 | ||||||
|---|---|---|---|---|---|---|---|---|
| 原始极大值 | 标准化极大绝对值 | 样本编号 | 海域 | 原始极小值 | 标准化极小绝对值 | 样本编号 | 海域 | |
| 胴长(ML) | 525 | 1 | 42 | 3 | 80 | 0.011236 | 144 | 1 |
| 上头盖长(UHL) | 39.56 | 1 | 42 | 3 | 5.44 | 0.001758 | 464 | 1 |
| 上脊突长(UCL) | 50.77 | 1 | 162 | 1 | 4.44 | 0.002806 | 146 | 1 |
| 上喙长(URL) | 12.92 | 1 | 30 | 3 | 1.77 | 0.000897 | 464 | 1 |
| 上侧壁长(ULWL) | 45.33 | 1 | 42 | 3 | 5.68 | 0.01488 | 154 | 1 |
| 上翼长(UWL) | 12.64 | 1 | 186 | 3 | 1.67 | 0.004558 | 144 | 1 |
| 下头盖长(LHL) | 11.63 | 1 | 200 | 3 | 1.66 | 0.003009 | 298 | 1 |
| 下脊突长(LCL) | 24.1 | 1 | 26 | 3 | 2.64 | 0.002796 | 184 | 1 |
| 下喙长(LRL) | 12.86 | 1 | 303 | 1 | 1.47 | 0.00439 | 464 | 1 |
| 下侧壁长(LLWL) | 35.67 | 1 | 42 | 3 | 4.62 | 0.00161 | 23 | 1 |
| 下翼长(LWL) | 22.1 | 1 | 42 | 3 | 2.25 | 0.012594 | 327 | 1 |
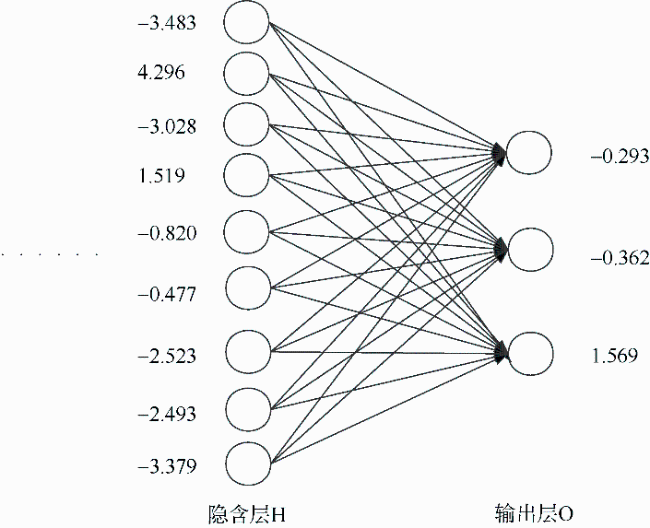
表2 I层和H层间的权重Tab. 2 The weights between input layer and hidden layer |
| I层 | H层 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | |
| ML | 0.51787 | 1.76012 | 1.17685 | 0.96851 | -0.62503 | -0.39972 | -2.30729 | -2.41620 | -1.21233 |
| UHL | 1.02004 | -0.11701 | 0.78299 | 0.97829 | 0.58829 | 1.06385 | -1.69839 | -0.67041 | 1.38657 |
| UCL | 0.20084 | 2.43943 | -0.47049 | -1.96293 | -0.06245 | -1.57966 | -0.29600 | 0.53659 | 0.31492 |
| URL | -0.04119 | 0.14562 | -0.99725 | -1.18086 | -1.72815 | -0.48685 | 1.37671 | -1.35249 | -1.31960 |
| ULWL | 1.04319 | -1.37696 | -2.54213 | 2.98592 | -1.81316 | 1.65405 | -1.53345 | -0.57482 | 0.30500 |
| UWL | -0.02106 | 1.29860 | 0.10095 | -1.31129 | -1.31384 | 0.01470 | -0.91160 | 0.56923 | 1.04699 |
| LHL | -1.62204 | 0.94419 | -1.77699 | 1.38963 | 0.65958 | -1.69024 | 0.19457 | 1.55272 | -0.54919 |
| LCL | 0.74875 | 0.65564 | 0.55693 | 0.92801 | -1.63957 | -1.29223 | 0.28282 | -0.22000 | 1.34480 |
| LRL | 0.61926 | -0.12646 | -1.45393 | 0.27165 | 0.40933 | -0.14337 | 0.97641 | -1.35238 | 1.06185 |
| LLWL | -1.90206 | 1.97220 | 0.89296 | 0.47994 | -1.69910 | -1.48343 | 0.13988 | 0.69561 | -1.31014 |
| LWL | 0.60720 | -0.28580 | -1.70620 | 1.63012 | -1.03251 | -0.77968 | 1.41782 | 0.98721 | -0.59789 |
表3 H层和O层间的权重Tab. 3 The weights between hidden layer and output layer |
| H层 | O层 | ||
|---|---|---|---|
| 西北印度洋 | 中东太平洋 | 南海 | |
| H1 | 0.16992 | -0.24264 | -0.10120 |
| H2 | -0.55120 | 2.30283 | -1.69967 |
| H3 | -0.30025 | 2.32027 | -2.03169 |
| H4 | 1.84776 | -1.90292 | 0.03078 |
| H5 | -0.15796 | 0.22273 | -0.12153 |
| H6 | 1.05078 | -1.17558 | 0.15734 |
| H7 | -0.47428 | -1.20609 | 1.73593 |
| H8 | -0.28751 | -1.01167 | 1.41477 |
| H9 | 0.44897 | 0.21886 | 0.99420 |
表4 动量自适应学习方法下训练集识别结果Tab. 4 The recognition results of the training set with the momentum adaptive learning method |
| 海区 | 样本总数 | 识别样本数 | 识别率 | ||
|---|---|---|---|---|---|
| 西北印度洋 | 中东太平洋 | 南海 | |||
| 西北印度洋 | 90 | 80 | 8 | 2 | 88.89% |
| 中东太平洋 | 189 | 2 | 170 | 17 | 89.95% |
| 南海 | 219 | 0 | 7 | 212 | 96.80% |
表5 动量自适应学习方法下测试集识别结果Tab. 5 The recognition results of the test set with the momentum adaptive learning method |
| 海区 | 样本总数 | 识别样本数 | 识别率 | ||
|---|---|---|---|---|---|
| 西北印度洋 | 中东太平洋 | 南海 | |||
| 西北印度洋 | 13 | 13 | 0 | 0 | 100% |
| 中东太平洋 | 27 | 0 | 24 | 3 | 88.89% |
| 南海 | 34 | 0 | 2 | 32 | 94.12% |
表6 各种BP学习方法的识别效果Tab. 6 The recognition results with different sorts of BPNN learning methods |
| 学习方法 | 各海区识别率 | 总识别率 | ||
|---|---|---|---|---|
| 西北印度洋 | 中东太平洋 | 南海 | ||
| 梯度下降法 | 46.15% | 70.37% | 88.24% | 74.32% |
| 单一动量法 | 53.85% | 70.37% | 91.12% | 77.03% |
| 单一自适应法 | 84.62% | 81.48% | 94.12% | 87.84% |
| 动量自适应法 | 100% | 88.89% | 94.12% | 93.24% |
| [1] |
陈芃, 方舟, 陈新军, 2015. 基于角质颚外部形态学的柔鱼种群判别[J]. 海洋渔业, 37(1):1-9.
|
| [2] |
陈艳, 2020. 基于BP神经网络的人脸识别方法分析[J]. 信息与电脑, 32(23):158-159.
|
| [3] |
关小芳, 2014. 动量自适应学习速率梯度下降法神经网络电力负荷预测[J]. 电气开关, 52(5):49-51.
|
| [4] |
李建华, 陈新军, 陈芃, 等, 2018. 赤道海域茎柔鱼角质颚形态及生长特征分析[J]. 海洋渔业, 40(4):385-393.
|
| [5] |
李敏, 张鹏, 张俊, 等, 2019. 南海鸢乌贼的遗传差异: 种群分化还是种间分化[J]. 中国水产科学, 26(1):133-140.
|
| [6] |
刘必林, 陈新军, 2009. 头足类角质颚的研究进展[J]. 水产学报, 33(1):157-164.
|
| [7] |
毛振宇, 李方利, 叶玉明, 等, 2019. 基于BP神经网络算法的电缆局部放电类型模式识别[J]. 机电信息, (27):20-22.
|
| [8] |
邱靖, 刘继荣, 曹志勇, 等, 2019. 基于卷积神经网络的水稻病害图像识别研究[J]. 云南农业大学学报(自然科学), 34(5):884-888.
|
| [9] |
王莉, 张紫烨, 郭晓东, 等, 2019. 基于粒子群优化BP神经网络的心电信号分类方法[J]. 自动化与仪表, 34(9):84-87.
|
| [10] |
王莉莉, 刘洪波, 陈德运, 等, 2018. 自适应与附加动量BP神经网络的ECT流型辨识[J]. 哈尔滨理工大学学报, 23(1):105-110.
|
| [11] |
王振华, 曲念毅, 钟元芾, 等, 2020. 一种改进深度卷积神经网络的海岛识别方法[J]. 上海海洋大学学报, 29(3):474-480.
|
| [12] |
徐杰, 刘尊雷, 李圣法, 等, 2016. 东海剑尖枪乌贼角质颚的外部形态及生长特性[J]. 海洋渔业, 38(3):245-253.
|
| [13] |
许嘉锦, 2003. Octopus与Cistpous属章鱼口器地标点之几何形态学研究[D]. 台北, 中国: 国立中山大学海洋生物研究所: 1-72.
|
| [14] |
叶斌, 雷燕, 2004. 关于BP网中隐含层层数及其节点数选取方法浅析[J]. 商丘职业技术学院学报, 3(6):52-53, 60.
|
| [15] |
袁曾任, 1999. 人工神经元网络及其应用[M]. 北京: 清华大学出版社.
|
| [16] |
张德丰, 2018. MATLAB R2017a人工智能算法[M]. 北京: 电子工业出版社. ZHANG DEFENG, 2018.
|
| [17] |
郑小东, 王如才, 刘维青, 2002. 华南沿海曼氏无针乌贼 Sepiella maindroni表型变异研究[J]. 青岛海洋大学学报, 32(5):713-719.
|
| [18] |
周开利, 康耀红, 2005. 神经网络模型及其matlab仿真程序设计[M]. 北京: 清华大学出版社.
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}