
Journal of Tropical Oceanography >
The framework, reservoir characteristics and reef formation model of Miocene algal reef dolomite in the Xisha Islands*
Copy editor: YIN Bo
Received date: 2023-02-21
Revised date: 2023-05-25
Online published: 2023-06-14
Supported by
National Key Research and Development Program(2017FY201407)
National Key Basic Research and Development Program(2012CB956004)
National Key Oil and Gas Project(2011ZX05025-002-04)
National Natural Science Foundation of China(41106064)
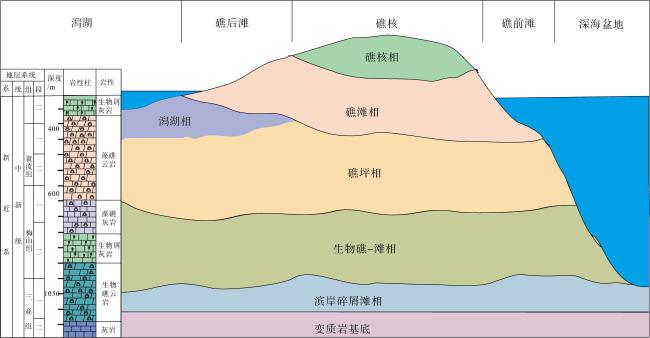
Traditional bioreef genesis theory either lacks evidences for algae forming strong bioreef anti-wave frameworks or attributes them to an accessory reef biotic community. In this paper, microfocal X-ray (X-CT) scanning imaging technology is used to study the core of the Aethesolithon nanhaiensis reef framework rocks and Halimeda reef proglottid rocks of Miocene age in Wells Xike 1 and Xiyong 2. Through three-dimensional tomography visual perspective images, it was found that the wave resistance structure of algal reef created by calcareous algal organisms, including typical Aethesolithon nanhaiensis reef framework structure, baffle-structure rocks and Halimeda reef proglottid structure. It also verifies that algal reefs are important Miocene oil-gas reservoirs in the South China Sea depositional basins by quantifying the throat, areal porosity and porosity parameters using threshold approach. Besides, the genetic model of algal reef is put forward: the reef-building calcium algal algae adapted to the sea level rise and fall gradually succession-replacement, and after calcification, burial and fossilization, dolomitization, sedimentary algal reef dolomitization, resulting in the growth of the longitudinal sequence of the Xisha Reef in Miocene.
CHEN Shu , XU Hong , LU Shushen , Zhang Haiyang , MA Yazeng , LUO Jinxiong . The framework, reservoir characteristics and reef formation model of Miocene algal reef dolomite in the Xisha Islands*[J]. Journal of Tropical Oceanography, 2024 , 43(2) : 140 -153 . DOI: 10.11978/2023022
图1 “西科1井”、“西永2井”藻礁白云岩微焦X光扫描分析样品a. 岩芯1, 仙掌藻礁白云岩, “西科1井”454.46m; b. 岩芯2, 南海奇石藻白云岩, “西科1井”547.38m; c. 岩芯3, “西永2井”550m; d. 岩芯4, “西永2井”400m; e. 岩芯5, “西永2井”520m; f. 岩芯6, 岩芯5的一小块 Fig. 1 Microfocal X-ray scanning samples of algal reef dolomite from Wells Xike 1 and Xiyong 2. (a) Core 1, dolomite of Halimeda Reef, Well Xike 1454.46 m; (b) Core 2, dolomite of Aethesolithon nanhaiensis, Well Xike 1547.38 m; (c) Core 3, Well Xiyong 2550 m; (d) Core 4, Well Xiyong 2400 m; (e) Core 5, Well Xiyong 2520 m; (f) core 6, a small piece of core 5 |
图6 “西科1井”岩心4切片a. 原始切片; b. 经过处理后的切片, 红色部分表示致密部分; c. 切片x、y、z轴3个方向的展示; d. 样品中最长一根藻类, 红色线条表示测量部位, 数字表示藻体的长度和宽度 Fig.6 Images of Core 4 sliced of Well Sike 1. (a) Original slice; (b) slice after treatment, the red spots indicate tight parts; (c) presentations of the slice along x, y, and z; (d) the longest alga among the samples. The red line indicates the measuring position. The numbers are the length and width of the algae |
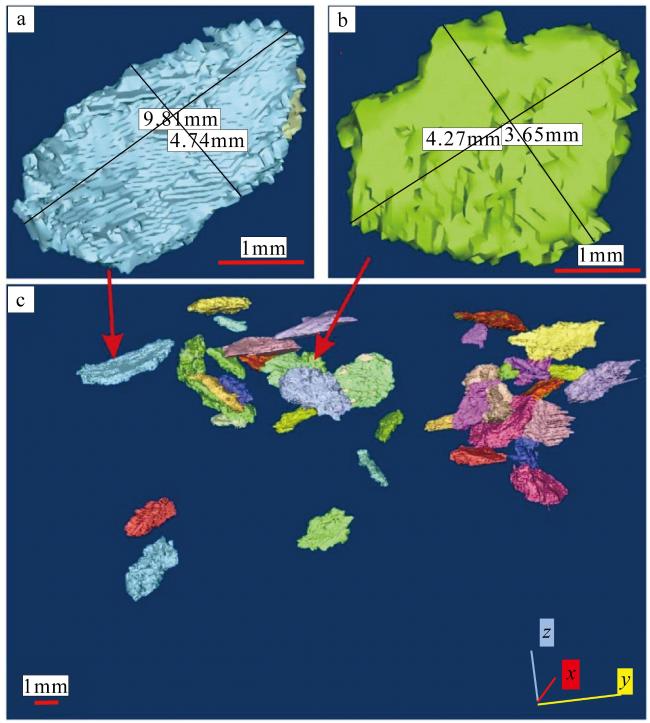
图7 “西科1井”仙掌藻化石三维空间表征a. 长9.81mm, 宽4.74mm的节片; b. 长4.27mm, 宽3.65mm的节片; c. 叠置在一起的节片。以不同颜色区分叠置在一起仙掌藻节片 Fig. 7 Spatial representation of the Halimeda fossils from XK-1. (a) Segments 9.81 mm long and 4.74 mm wide; (b) Segments 4.27 mm long and 3.65 mm wide; (c) Segments stacked together. Superimposed together in different colors |
图8 “西科1井”南海奇石藻三维重构图a、b为整体重构图; c、d为珊瑚藻重构图。绿色部分为南海奇石藻藻体; 紫色部分为基质 Fig. 8 3D reconstructions of the Aethesolithon nanhaiensis from XK-1. (a) and (b) are the whole reconstructions; (c) and (d) are reconstructions of the coralline algae. The green part is the algal body of Aethesolithon nanhaiensis; The purple part is the matrix |
图9 “西永2井”岩心4三维重构图a. 高精度CT重构后的岩石样品表面; b. 透析后藻类在样品中的分布; c. 可区分出碳酸盐岩中的重矿物; d. 高精度CT扫描断层横切面 Fig. 9 Three-dimensional recomposition of core 4 in Well 2 of Xiyong. (a) Rock sample surface after high-precision CT reconstruction; (b) Distribution of dialyzed algae in the sample; (c) heavy minerals in the carbonatite can be distinguished; (d) Cross section of high-precision CT scan tomography |
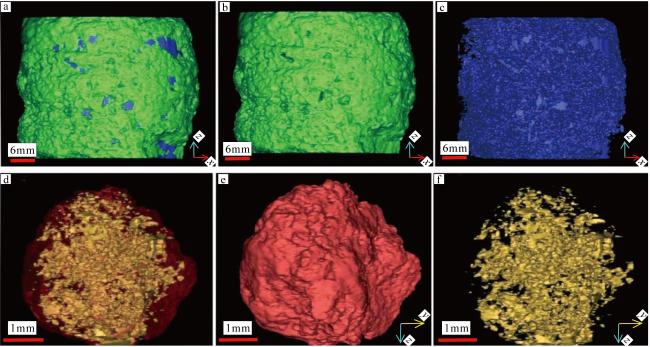
图10 岩芯5、6的三维重构图a. 岩芯5的整体重构图; b. 岩芯5的骨架重构图; c. 岩芯5的孔隙重构图; d. 岩芯6的整体重构图; e. 岩芯6的骨架重构图; f. 岩芯6的孔隙重构图; 绿色为基质, 蓝色为孔隙; 红色为基质, 黄色为孔隙 Fig. 10 3D reconstructions of cores 5 and 6. (a), (b) and (c) are the whole reconstruction, core skeleton reconstruction and pore reconstruction of core 5, respectively; (d), (e) and (f) are the whole reconstruction, core skeleton reconstruction and pore reconstruction of core 6, respectively; Green is the matrix, blue is the pores, red is the matrix and yellow is the pores |
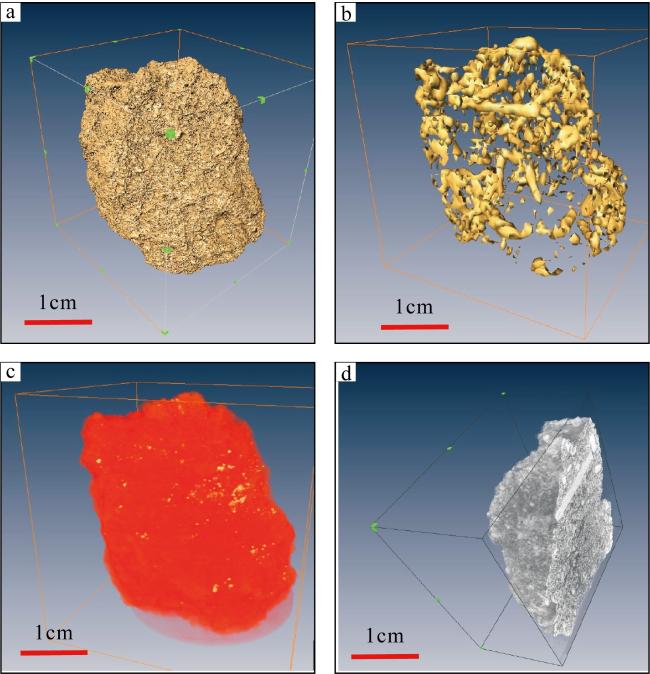
图11 “西科1井”岩心1孔隙三维重构图a. z轴距第一张切片d=1.32mm的切片; b. 岩芯三维透视图; c. 图a切片孔隙展示; d. 图b岩芯孔隙三维重构, 图中黑色实线分别指示长、宽、高, 长为25mm, 宽为13.19mm, 高为16.19mm Fig. 11 3D pore reconstruction of the core1 from XK-1. (a) Slice along z at d = 1.32 mm from the first slice; (b) 3D perspective of the core; (c) pore presentation of slice a; (d) 3D pore reconstruction of core b. The solid black lines in the figure indicate the length, width, and height respectively. The length is 25 mm, the width is 13.19 mm, and the height is 16.19 mm |
图12 岩芯4、岩芯5不同方向扫描图片的二值化处理后图像a, b, c为岩芯4的x, y, z轴切片, 红色部分为颗粒和基质, 黄色部分为孔隙; d为重构图像; e, f, g为岩芯5的x, y, z轴切片, 绿色部分为颗粒和基质, 蓝色部分为孔隙; h为重构图像 Fig. 12 Binarized scan images of core 4 and core 5 in different directions. (a), (b) and (c) are slices along x, y, z of core 4, the red spots indicate grains and matrixes, the yellow spots indicate pores; (d) reconstruction image; (e), (f) and (g) are slices along x, y, z of core 5, the green spots indicate grains and matrixes, the blue spots indicate pores; (h) reconstruction image |
表1 岩芯4、5、6孔隙结构参数的CT测量结果Tab. 1 CT measurements of pore structure parameters of cores 4, 5 and 6 |
| 样品 | 孔隙表面积/mm2 | 孔隙体积/mm3 | 岩石骨架体积/mm3 | 总体积/mm3 | 孔隙度/% | 平均面孔率/% |
|---|---|---|---|---|---|---|
| 岩芯4 | 172500.80 | 18649.50 | 66521.56 | 85171.06 | 21.90 | 26 |
| 岩芯5 | 121500.00 | 12778.40 | 88688.95 | 101467.30 | 12.59 | 19 |
| 岩芯6 | 97.08 | 2.12 | 18.41 | 20.52 | 10.32 | 21 |
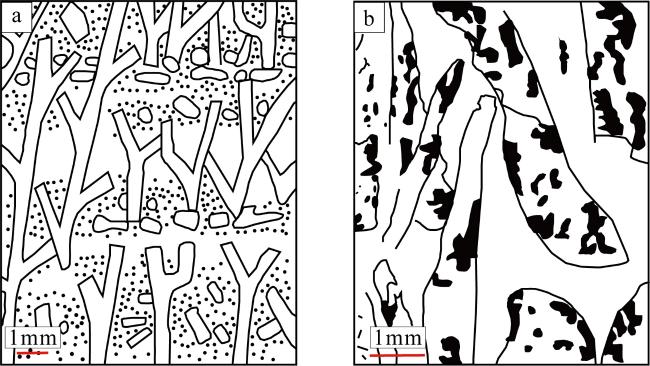
图13 两类钙藻岩石结构及其岩石类型图[据许红等(1999)修改]a. 南海奇石藻格架-障积结构(绘图); b. 仙掌藻节片结构(偏光显微镜照片) Fig. 13 Images showing the rock structures and rock types of the two calcareous algae (Xu, 1999, with modification). (a) Aethesolithon nanhaiensis framework-baffle structure (plot); (b) Halimeda proglottid structure (polarized photomicrograph) |
| [1] |
白斌, 朱如凯, 吴松涛, 等, 2013. 利用多尺度CT成像表征致密砂岩微观孔喉结构[J]. 石油勘探与开发, 40(3): 329-333.
|
| [2] |
陈斯忠, 胡平忠, 1987. 珠江口盆地第三纪生物礁及其找油意义[J]. 中国海上油气, 1(1): 3-10.
|
| [3] |
陈浩, 黄继新, 常广发, 等, 2018. 基于全岩心CT的遗迹化石识别及沉积环境分析: 以加拿大麦凯Ⅲ油砂区块为例[J]. 古地理学报, 20(4): 703-712.
|
| [4] |
范嘉松, 1996. 中国生物礁与油气[M]. 北京: 科学出版社.
|
| [5] |
高林志, 邢裕盛, 刘桂芝, 1995. 吉林浑江地区新元古代微古植物群及其沉积环境[J]. 地层古生物论文集, (2): 1-23, 124-127.
|
| [6] |
侯健, 李振泉, 张顺康, 等, 2008. 岩石三维网络模型构建的实验和模拟研究[J]. 中国科学, 8(11): 1563-1575.
|
| [7] |
蒋观利, 吴青柏, 蒲毅彬, 等, 2005. 甲烷水合物形成过程的CT识别原理和成像特征[J]. 天然气地球科学, 16(6): 814-817.
|
| [8] |
李菊花, 郑斌, 2015. 微观孔隙分形表征新方法及其在页岩储层中的应用[J]. 天然气工业, 35(5): 52-59.
|
| [9] |
李玉彬, 李向良, 张奎祥, 等, 2000. 微焦点X射线计算机层析(CMT)及其在石油研究领域的应用[J]. CT理论与应用研究, 9(3): 35-40.
|
| [10] |
李越, 穆西南,
|
| [11] |
李越, 穆西南, 袁金良, 等, 2001. 山东中寒武世钙藻礁的层位拟定[C]. 中国古生物学会第21届学术年会论文摘要集: 20-21.
|
| [12] |
廉培庆, 高文彬, 汤翔, 等, 2020. 基于CT扫描图像的碳酸盐岩油藏孔隙分类方法[J]. 石油与天然气地质, 41(4): 852-861.
|
| [13] |
刘月田, 郭分乔, 涂彬, 等, 2005. 全岩心非均匀径向渗流各向异性渗透率测定方法[J]. 石油学报, 26(6): 66-68.
|
| [14] |
刘志礼, 1990. 化石藻类学导论[M]. 北京: 高等教育出版社.
|
| [15] |
马文国, 刘傲雄, 2011. CT扫描技术对岩石孔隙结构的研究[J]. 中外能源, 16(7): 54-56.
|
| [16] |
马骁, 许红, 付和平, 等, 2021. 海洋造礁仙掌藻研究进展及西沙石岛仙掌藻[J]. 海洋地质前沿, 37(6): 77-83.
|
| [17] |
穆西南, 1982. 西藏的一些钙藻化石. 青藏高原科学考察丛书, 西藏古生物(第五分册)[M]. 北京: 科学出版社:205-240.
|
| [18] |
穆西南, 王玉净, 1985. 西藏定日始新世的一些钙藻化石[J]. 微体古生物学报, (3): 289- 299, 323-325.
|
| [19] |
穆西南, 严惠君, 李越, 等, 2003b. 华北地台东部中寒武世微生物礁的时空分布[J]. 微体古生物学报, (3): 279-285.
|
| [20] |
穆西南, 袁训来, 2003a. 绪论: 化石藻类和叠层石[J]. 微体古生物学报, (1): 1-2.
|
| [21] |
彭瑞东, 杨彦从, 鞠杨, 等, 2011. 基于灰度CT图像的岩石孔隙分形维数计算[J]. 科学通报, 56(26): 2256-2266.
|
| [22] |
钱凯, 王淑芬, 1986. 济阳坳陷下第三系礁灰岩及礁灰岩油气藏[J]. 石油勘探与开发, (5): 1-7.
|
| [23] |
钱凯, 王素民, 刘淑范, 等, 1980. 华东北部下第三系礁灰岩的发现及其石油地质意义[J]. 科学通报, 25(24): 1140-1142.
|
| [24] |
孙海, 姚军, 张磊, 等, 2014. 基于孔隙结构的页岩渗透率计算方法[J]. 中国石油大学学报: 自然科学版, 38(2): 92-98.
|
| [25] |
孙建孟, 闫国亮, 姜黎明, 等, 2014. 基于数字岩心研究流体性质对裂缝性低渗透储层弹性参数的影响规律[J]. 中国石油大学学报:自然科学版, 38(3): 39-44.
|
| [26] |
王玉净, 1976. 珠穆朗玛峰地区晚白垩世及早第三纪钙藻化石. 珠穆朗玛峰地区科学考察报告(1966—1968), 古生物(第二分册)[M]. 北京: 科学出版社:425-457.
|
| [27] |
王玉净, 勾韵娴, 章炳高, 等, 1996. 西沙群岛西琛一井中新世地层、古生物群和古环境研究[J]. 微体古生物学报, (3): 215-223.
|
| [28] |
吴洁, 刘成东, 张守鹏, 等, 2012. 显微CT技术在石油地质中的应用初探[J]. 江西科学, 30(5): 634-637.
|
| [29] |
邢裕盛, 1987. 前寒武纪微古植物学在我国的发展和应用[J]. 地层古生物论文集, (1): 8-28 (in Chinese).
|
| [30] |
邢裕盛, 刘桂芝, 1973. 燕辽地区震旦纪微古植物群及其地质意义[J]. 地质学报, (1): 1-64 (in Chinese).
|
| [31] |
邢裕盛, 刘桂芝, 1980. 鄂西震旦亚界微古植物群及其地层意义[J]. 地层古生物论文集, (1): 1-14, 91-96, 124.
|
| [32] |
邢裕盛, 刘桂芝, 1982. 中国晚前寒武纪微古植物群及其地层意义[C]. 中国地质科学院文集, (4): 55-67.
|
| [33] |
许红, 1992. 中国海域及邻区含油气盆地生物礁的对比研究[J]. 海洋地质与第四纪地质, 12(4): 41-52.
|
| [34] |
许红, 蔡峰, 王玉净, 等, 1999. 西沙中新世生物礁演化与藻类的造礁作用[J]. 科学通报, 43(13): 1435-1439 (in Chinese).
|
| [35] |
姚艳斌, 刘大锰, 蔡益栋, 等, 2010. 基于NMR和X-CT的煤的孔裂隙精细定量表征[J]. 中国科学: 地球科学, 40(11): 1598-1607 (in Chinese).
|
| [36] |
曾鼎乾, 1981. 南海北部大陆架第三系[M]. 广州: 广东科技出版社.
|
| [37] |
章海宁, 姜黎明, 张金功, 等, 2014. 岩石结构对碎屑岩储层弹性参数影响的数值研究[J]. 兰州大学学报: 自然科学版, 50(6): 773-778.
|
| [38] |
张明书, 何起祥, 业治铮, 1989. 西沙生物礁碳酸盐沉积地质学研究[M]. 北京: 科学出版社: 1-117.
|
| [39] |
赵永刚, 赵明华, 赵永鹏, 等, 2006. 一种分析碳酸盐岩孔隙系统数字图像的新方法[J]. 天然气工业, 26(12): 75-78.
|
| [40] |
朱浩然, 1979. 山东滨县下第三系沙河街组的藻类化石[J]. 古生物学, 18(4): 327-346.
|
| [41] |
朱袁智, 沙庆安, 郭丽芬, 等, 1997. 南沙群岛永暑礁新生代珊瑚礁地质[M]. 北京: 科学出版社.
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
/
| 〈 |
|
〉 |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}