
Journal of Tropical Oceanography >
Ovarian development, oocyte and yolk production of Paphia textile in Beibu Gulf, Guangxi
Copy editor: YIN Bo
Received date: 2023-05-30
Revised date: 2023-07-14
Online published: 2023-07-25
Supported by
Guangxi Innovation Driven Development Project(Guike AA19254032)
In this study, ovarian development, reproductive cycle, egg development and its ultrastructure of Paphia textile in Beibu Gulf of Guangxi were investigated using anatomical observation, ovarian tissue sectioning and transmission electron microscopy techniques. The results showed that P. textile has one reproductive cycle in one year, and the ovarian development of P. textile can be divided into five stages: proliferating stage (May to July), growing stage (July to September), maturing stage (September to November), spawning stage (November to January of the next year), and resting stage (January to April of the next year). According to the relevant characteristics of the egg development and vitellogenesis of P. textile, it can be divided into four main stages: the first stage is the oocytes stage, the second stage is the phase of previtellogenesis oocytes stage, the third stage is the phase of vitellogenesis oocytes stage, and the fourth stage is the phase of postvitellogenic oocytes stages. The phase of vitellogenesis is an important stage of vitellogenesis, during which mitochondria, endoplasmic reticulum, golgiosome, lysosomes and other organelles form yolk granule in different ways. The water temperature in the sea area was closely related to the ovarian development of P. textile and was an important factor affecting the ovarian development of P. textile. P. textile is hermaphroditic, and no hermaphroditism has been found. The results of the study provide providing basic data for reproductive biology, artificial breeding, breeding and germplasm resources protection of P. textile.
XU Bingjie , LIU Yiming , LIAN Changpeng , WU Tao , PAN Ying . Ovarian development, oocyte and yolk production of Paphia textile in Beibu Gulf, Guangxi[J]. Journal of Tropical Oceanography, 2024 , 43(2) : 48 -58 . DOI: 10.11978/2023070
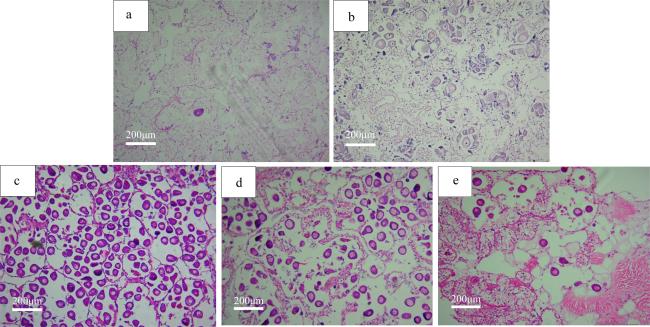
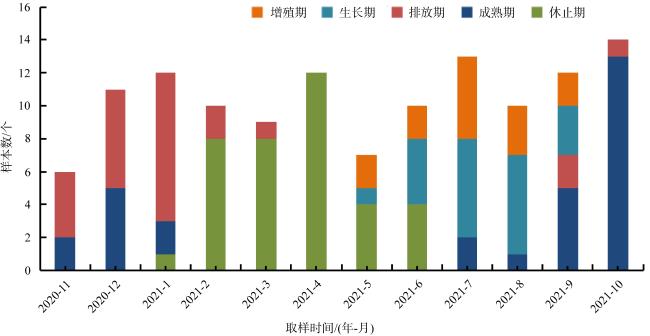
表1 广西北部湾海域织锦巴非蛤卵巢发育各期的周年分布Tab. 1 Annual distribution of ovarian development stages of P. textile in sea area of Beibu Gulf, Guangxi |
| 取样日期/(年-月-日) | 卵巢发育各期内的个体数 | 总数/个 | 盐度/‰ | 水温/℃ | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | |||||
| 2020-11-29 | 2 | 4 | 6 | 27.4 | 22.0 | ||||
| 2020-12-21 | 5 | 6 | 11 | 27.5 | 17.2 | ||||
| 2021-01-08 | 2 | 9 | 1 | 12 | 29.4 | 15.6 | |||
| 2021-02-24 | 2 | 8 | 10 | 30.9 | 17.3 | ||||
| 2021-03-18 | 1 | 8 | 9 | 30.0 | 21.0 | ||||
| 2021-04-22 | 12 | 12 | 30.0 | 27.5 | |||||
| 2021-05-20 | 2 | 1 | 4 | 7 | 28.0 | 29.5 | |||
| 2021-06-23 | 2 | 4 | 4 | 10 | 28.4 | 30.5 | |||
| 2021-07-15 | 5 | 6 | 2 | 13 | 29.0 | 29.7 | |||
| 2021-08-18 | 3 | 6 | 1 | 10 | 30.0 | 29.8 | |||
| 2021-09-17 | 2 | 3 | 5 | 2 | 12 | 31.0 | 29.8 | ||
| 2021-10-20 | 13 | 1 | 14 | 29.0 | 25.5 | ||||
表2 织锦巴非蛤各期卵母细胞特征Tab. 2 The characteristics of oocytes at different stages of P. textile |
| 时期 | 细胞特点 | 细胞器特点 | 卵黄特点 |
|---|---|---|---|
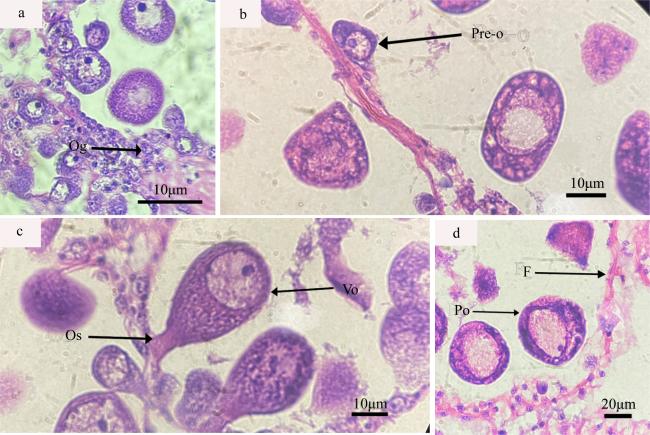
| 卵原细胞期 | 卵原细胞紧靠滤泡壁, 由泡壁上的生殖上皮细胞增殖而来, 形状呈椭圆形或圆形。细胞卵径约为0.94~8.96μm, 细胞间界限不清晰, 但可观察到细胞核。 | 细胞核内核质聚集, 核仁明显, 细胞在此期整体染色较深。电镜下可观察到卵原细胞的胞质呈颗粒状, 胞质存在较多的囊泡, 线粒体等细胞器较少, 核仁不位于细胞核的中央。 | 此期未开始卵黄合成。 |
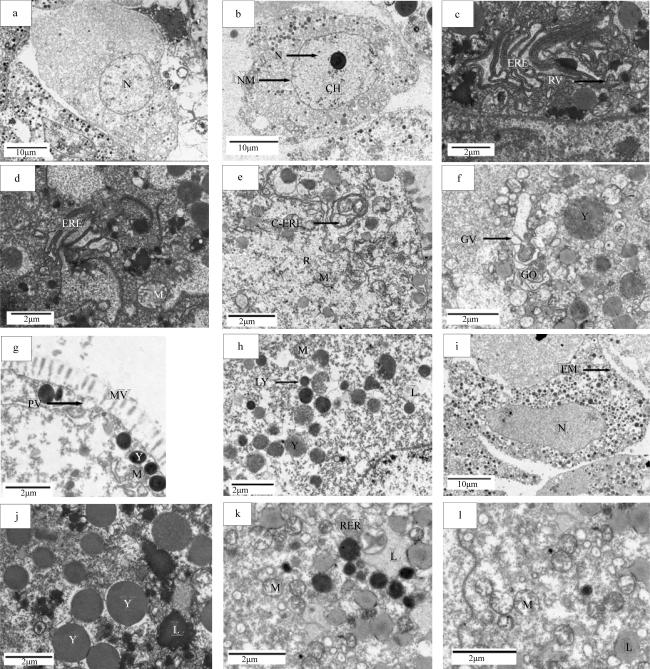
| 卵黄合成前期 | 在滤泡内壁周围。细胞卵径约为9.51~15.43μm, 前期与滤泡壁相贴合, 后期逐渐朝向滤泡中心, 可见卵质有一部分为变窄狭长, 变为滤泡壁相连的卵柄。形状为椭圆形或梨形。细胞核的染色质有聚集成团块的趋势, 同时卵母细胞胞质内存在许多大小、形状不一的囊泡。 | 细胞核中核质膨胀成泡状, 逐渐透亮, 形成一个生发泡, 卵母细胞胞质中可观察到线粒体等细胞器, 高尔基体复合体开始形成。粗面内质网形成少量囊泡。脂滴的数量开始增多。 | 此时有少量卵黄粒在合成, 卵黄粒无膜结构, 体积较小, 呈球状。同时胞质周围有大量的糖原颗粒分布。 |
| 卵黄合成期 | 在滤泡周围和靠中央位置, 细胞卵径约15.06~57.20μm, 细胞形态从扁平或不规则逐渐变为梨形或者葡萄状, 卵柄变窄, 与滤泡壁脱离或仅剩一点柄状连接。细胞核偏向一边, 细胞内卵黄粒增多。卵母细胞表面微绒毛发达。卵母细胞胞质内有大量内质网、线粒体等细胞器。 | 卵母细胞中细胞器数量多。内质网发达, 分为层状和环状结构, 环状结构还会包围脂滴进行卵黄粒的合成。高尔基体复合体在边缘形成囊泡, 包囊包裹线粒体等进行卵黄粒沉淀。 | 此时卵黄大量合成, 卵黄的来源多种, 形状与大小不一, 不同来源的卵黄粒沉积程度有差别。 |
| 卵黄合成后期 | 成熟卵子形状为球形, 卵子间相互挤压, 分布在滤泡中央位置。卵径比大生长期的卵母细胞稍大一些, 细胞卵径约为 48.78~62.54μm, 呈饱满状, 核膜清晰, 核仁靠在细胞核的一边、卵黄粒多, 大量分布在核质中。胞质内充满卵黄粒, 卵黄饱满, 成熟卵细胞相互挤压, 细胞核清晰、有少部分可观察到存在2个核仁。 | 卵母细胞的细胞器数量大幅减少。内质网和高尔基体复合体等细胞器特征已经不明显, 形成包囊。Ⅱ型线粒体内部脊消失, 出现空泡化, 合成卵黄粒。 | 大量卵黄分布在成熟卵子内, 多为球形, 大部分发育成熟。卵黄粒在卵母细胞内呈现饱满状, 数量多、体积大。 |
图3 织锦巴非蛤各期雌性生殖细胞a. 卵原细胞期, Og: 卵原细胞; b. 卵黄合成前期卵母细胞期, Pre-o:卵黄合成前期卵母细胞; c. 卵黄合成期卵母细胞期, Os: 卵柄, Vo: 卵黄合成期卵母细胞; d. 卵黄合成后期卵母细胞期, Po: 卵黄合成后期卵母细胞, F: 滤泡壁 Fig. 3 The different stages of female reproductive cell of P. textile. (a) Ogonium period, Og: Oogonium; (b) Period of previtollogenic, Pre-o: Previtellogenic stage oocyte; (c) Period of vitellogenesis oocyte, Vo: Vitellogenic stage oocyte, Os: Oocyte stalk; (d) Period of postvitellogenic stage, Po: Postvitellogenic stage oocyte, F: Follicular wall |
图4 织锦巴非蛤卵母细胞发育的超微结构观察a. 卵黄合成前期卵母细胞, 示细胞核(N); b. 卵黄合成期卵母细胞, 示细胞核(N)、核膜(NM)、核质(CH); c. 卵黄合成期, 示粗面内质网(ERE)、内质网囊泡(RV); d. 卵黄合成期, 示粗面内质网(ERE); e. 卵黄合成期, 示圆圈状粗面内质网(C-ERE)、核糖体(R)、线粒体(M); f. 卵黄合成期, 示高尔基体复合体(GO)、高尔基体囊泡(GV)、卵黄(Y); g. 卵黄合成期, 示微绒毛(MV)、吞噬小泡(PV); h. 卵黄合成期, 示脂滴(L)、溶酶体(LY)、卵黄(Y)、线粒体(M); i. 卵黄合成后期卵母细胞, 示细胞膜(EM)、细胞核(N); j. 卵黄合成后期, 示卵黄(Y)、脂滴(L); k. 卵黄合成后期, 示内质网包围脂滴(L)、粗面内质网(RER)、线粒体(M); l. 卵黄合成后期, 示线粒体空泡化(M)、脂滴(L) Fig. 4 Ultrastructural observation on the development of oocytes in P. textile. (a) The early period of vitellogenesis oocyte, showing nucleus (N); (b) The period of vitellogenesis oocyte, showing nucleus (N), nuclear membrane (NM) and chromatin (CH); (c) At vitellogenic stage, showing rough endoplasmic reticulum (RER) and rough endoplasmic reticulum vesicles (RV); (d) At vitellogenic stage, showing rough endoplasmic reticulum (RER); (e) At vitellogenic stage, showing circled rough endoplasmic reticulum (C-RER), ribosome (R), mitochondria (M); (f) At vitellogenic stage, showing Golgi complex (GO) and Golgi vesicle (GV), yolk granules (Y); (g) At vitellogenic stage, showing microvilli (MV) and pinocytosis vesicles (PV); (h) At vitellogenic stage, showing lipid droplet (L) and lysosome (LY), yolk granules (Y), mitochondria (M); (i) The late period of vitellogenesis oocyte, showing cell membrane (EM), nucleus (N); (j) At postvitellogenic stage, showing yolk granules (Y) and lipid droplet (L); (k) At postvitellogenic stage, showing Endoplasmic reticulum surrounds lipid droplets (L), rough endoplasmic reticulum (RER), mitochondria (M); (l) At postvitellogenic stage, showing cavitation of mitochondria (M), lipid droplet (L) |
| [1] |
白胡木吉力图, 马汝河, 高悦勉, 等, 2008. 大连海区青蛤的性腺发育和生殖周期[J]. 大连水产学院学报, 23(3): 196-199.
|
| [2] |
曹伏君, 刘永, 张春芳, 等, 2012. 施氏獭蛤(Lutraria sieboldii)性腺发育和生殖周期的研究[J]. 海洋与湖沼, 43(5): 976-982.
|
| [3] |
邓正华, 翟子钦, 魏海军, 等, 2022. 织锦巴非蛤幼虫对不同种类单胞藻的摄食和消化效果[J]. 南方农业学报, 53(5): 1448-1556.
|
| [4] |
顾海龙, 林志华, 沈伟良, 等, 2013. 泥蚶初级卵母细胞发育及卵黄发生的超微结构研究[J]. 海洋科学, 37(1): 49-53.
|
| [5] |
柯巧珍, 李琪, 闫红伟, 等, 2012. 山东北部沿海四角蛤蜊性腺发育年周期研究[J]. 中国海洋大学学报(自然科学版), 42(11): 28-34.
|
| [6] |
赖胜琪, 尹聪, 邱炬维, 等, 2022. 北部湾沿海织锦巴非蛤不同地理群体形态差异研究[J]. 广东农业科学, 49(7): 105-112.
|
| [7] |
李庆昌, 刘坦, 陈小明, 等, 2016. 织锦巴非蛤斧足颜色与总类胡萝卜素含量相关分析[J]. 海洋科学, 40(10): 120-125.
|
| [8] |
李霞, 梁峻, 闫喜武, 等, 2007. 人工控温促熟条件下不同海域菲律宾蛤仔的性腺发育[J]. 水产学报, 31(6): 853-859.
|
| [9] |
栗志民, 刘志刚, 韩伟贤, 2011. 织锦巴非蛤稚贝盐度适应性研究[J]. 海洋科学, 35(10): 96-102.
|
| [10] |
连昌朋, 吴韬, 王超奇, 等, 2022. 广西北海营盘海域钝缀锦蛤(Tapes conspersus)卵巢发育、卵子和卵黄发生的研究[J]. 热带海洋学报, 41(5): 170-179.
|
| [11] |
廖承义, 徐应馥, 王远隆, 等, 1983. 栉孔扇贝的生殖周期[J]. 水产学报, 7(1): 1-13.
|
| [12] |
刘海娟, 陈瑞芳, 曾梦清, 等, 2022. 织锦巴非蛤人工养殖技术研究[J]. 科学养鱼, 38(11): 61-63.
|
| [13] |
刘相全, 方建光, 包振民, 等, 2007. 中国蛤蜊繁殖生物学的初步研究[J]. 中国海洋大学学报(自然科学版), 37(1): 89-92.
|
| [14] |
吕敏, 李琪, 2022. 山东田横岛海域长牡蛎性腺发育及生化成分周年变化[J]. 中国海洋大学学报(自然科学版), 52(2): 33-40.
|
| [15] |
宁军号, 常亚青, 宋坚, 等, 2015. 偏顶蛤的性腺发育和生殖周期[J]. 中国水产科学, 22(3): 469-477.
|
| [16] |
上官步敏, 刘正琮, 1995. 缢蛏卵母细胞卵黄发生过程的超微结构研究[J]. 海洋与湖沼, 26(1): 48-52.
|
| [17] |
闻海波, 孙光兴, 丁图强, 等, 2020. 淮河橄榄蛏蚌繁殖类型与性腺发育观察[J]. 中国水产科学, 27(10): 1156-1160.
|
| [18] |
吴加莹, 戴明姝, 刘志刚, 等, 2023. 温度对织锦巴非蛤稚贝生存和生长的影响[J]. 南方水产科学, 19(2): 62-69.
|
| [19] |
吴明灿, 张立, 潘英, 等, 2015. 糙海参卵子发生及卵黄发生的超微结构[J]. 热带海洋学报, 34(3): 68-74.
|
| [20] |
巫旗生, 文宇, 曾志南, 等, 2017. 钝缀锦蛤繁殖周期和胚胎发育[J]. 中国水产科学, 24(3): 488-496.
|
| [21] |
徐凤山, 张素萍, 2008. 中国海产双壳类图志[M]. 北京: 科学出版社:244- 245.
|
| [22] |
杨林, 李琪, 闫红伟, 等, 2010. 山东沿海中国蛤蜊的繁殖生物学特性[J]. 中国水产科学, 17(3): 514-520.
|
| [23] |
应雪萍, 杨万喜, 2001. 泥螺卵黄发生过程中线粒体的变化[J]. 动物学研究, 22(5): 379-382.
|
| [24] |
曾庆民, 赵虹博, 邓素贞, 等, 2022. 织锦巴非蛤不同颜色斧足的转录组[J]. 水产学报, 46(9): 1562-1571.
|
| [25] |
翟子钦, 喻达辉, 白丽蓉, 2022. 织锦巴非蛤形态性状对体质量的影响[J]. 广东农业科学, 49(7): 113-119.
|
| [26] |
赵雯, 车宗豪, 刘括, 等, 2022. 日本海神蛤(Panopea japonica)性腺发育生物学零度及有效积温研究[J]. 海洋科学, 46(10): 78-84.
|
| [27] |
朱星海, 孙红振, 杨祖晶, 等, 2019. 风信标扇贝的性腺发育与繁殖周期规律研究[J]. 中国海洋大学学报(自然科学版), 49(2): 52-58.
|
| [28] |
庄启谦, 2001. 中国动物志: 软体动物门、双壳纲、帘蛤科[M]. 北京: 科学出版社:53- 54.
|
| [29] |
邹杰, 彭慧婧, 杨家林, 2019. 织锦巴非蛤人工种苗培育及浅海养殖试验[J]. 科学养鱼, 35(10): 57-58.
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}