
Journal of Tropical Oceanography >
Zooplankton community in the coastal waters of eastern Guangdong under the influence of human activities and ocean currents
Copy editor: SUN Cuici
Received date: 2023-07-30
Revised date: 2023-09-02
Online published: 2023-10-20
Supported by
Special Fund for Science and Technology Planning Project of Guangdong Province of China(2021B1212050023)
Special Fund for Science and Technology Planning Project of Guangdong Province of China(2023B1212060047)
National Natural Science Foundation of China(32171548)
Guangdong Basic and Applied Basic Research Foundation(2022A1515010656)
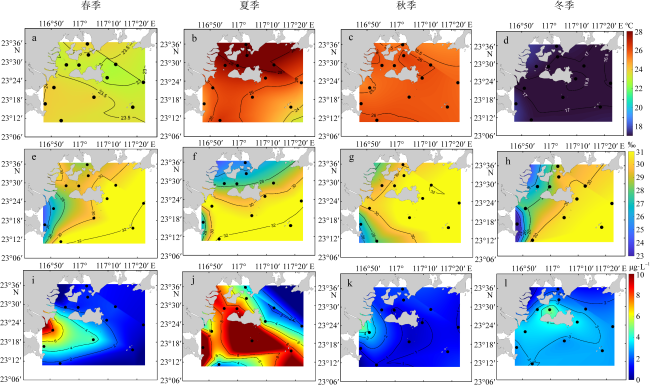
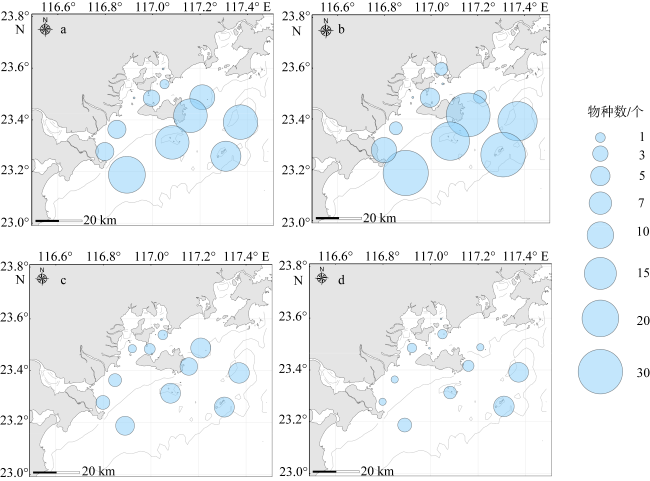
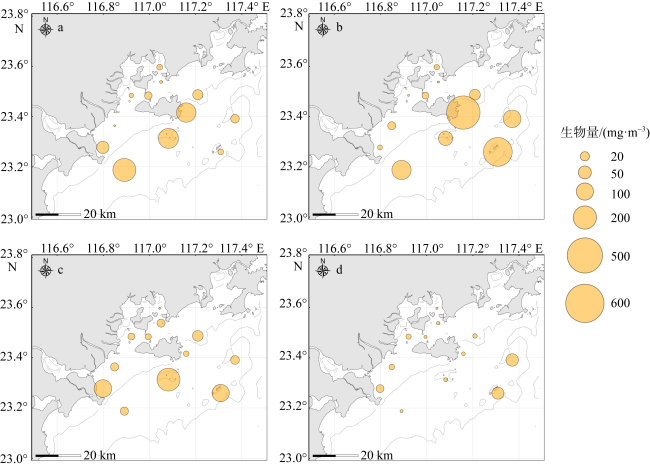
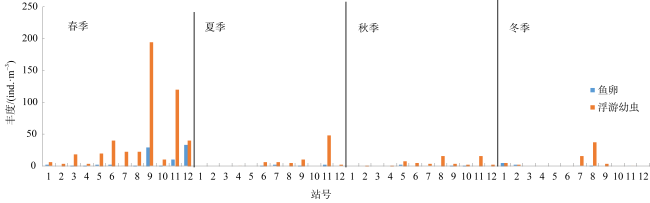
There are rich fishery resources and complicated hydrologic environment in the coastal waters of eastern Guangdong. In order to systematically improve the level of oceanic observation and the knowledge of ecological dynamic processes in the coastal waters of eastern Guangdong under the changing climate, a comprehensive investigation of ecological and environmental factors such as hydrodynamics, biology and chemistry during four seasons was carried out in May (spring), July (summer), October (autumn) 2022 and January 2023 (winter). Based on the taxonomic identification of zooplankton, the characteristics of community structure of zooplankton in the surveyed area were analyzed, and the effects of environmental factors on their spatial and temporal distribution were discussed. According to the environmental parameters, the survey area can be divided into the near-shore (mainly affected by human activities) and off-shore (mainly affected by ocean currents) areas, and the zooplankton were characterized by the seasonal and regional pattern. Seasonally, the species, biomass and abundance of zooplankton in spring and summer were higher than those in autumn and winter. Regionally, the species, biomass and abundance of zooplankton in the off-shore waters are generally higher than those in the near-shore waters. Moreover, there are differences in the contribution groups and dominant species to the total zooplankton abundance in different seasons in the off-shore waters. Although copepods and chaetognaths were the main groups of zooplankton, the abundance of planktonic larvae and fish eggs in spring was higher than in other seasons, and the peak value is concentrated in the southwestern Hanjiang Estuary, indicating that this surveyed area was still an important spawning ground in the coastal waters of eastern Guangdong. The increase in copepods species and abundance in summer is related to the high chlorophyll a concentration improved by coastal upwelling in eastern Guangdong. Warm-temperate species carried by Fujian and Zhejiang coastal currents in winter are the main contributors to zooplankton abundance. Although chlorophyll a concentrations are higher in the near-shore waters, zooplankton species, biomass and abundance are lower. Under the effect of anthropogenic activities and climate change, long-term systematic monitoring and analysis of the response of zooplankton to the dynamics of the marine environment are needed in the coastal waters of eastern Guangdong.
LIU Yuan , KE Zhixin , LI Kaizhi , TAN Yehui , LIANG Junce , ZHOU Weihua . Zooplankton community in the coastal waters of eastern Guangdong under the influence of human activities and ocean currents[J]. Journal of Tropical Oceanography, 2024 , 43(4) : 98 -111 . DOI: 10.11978/2023107
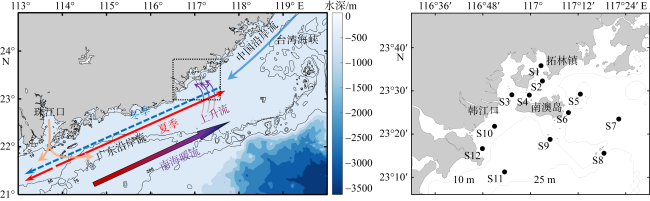
图1 粤东近海海流示意图(左图)和2022年5月—2023年1月四个季节浮游动物调查站位图(右图)该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2019)1825号的标准地图制作, 下同。左图虚线框为采样大致范围 Fig. 1 Schematic diagram of currents in the coastal waters of eastern Guangdong (left) and sampling stations of zooplankton in spring, summer, autumn and winter from May 2022 to January 2023 (right) |
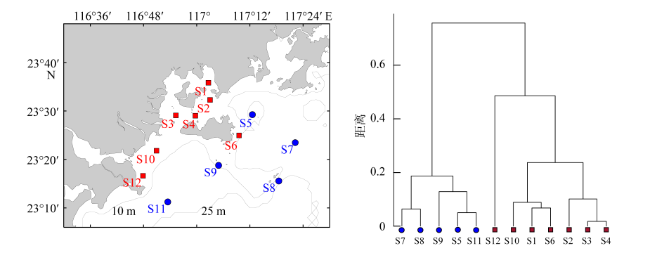
图3 2022年5月—2023年1月粤东近海表层环境因子聚类的调查站位分布(a)和聚类结果(b)■为主要受人类活动影响的站位, ●为主要受海流影响的站位 Fig. 3 Distribution of sampling stations based on surface environmental parameters clustering (a) and clustering results (b) in coastal waters of eastern Guangdong from May 2022 to January 2023 ■ represents stations mainly affected by human activities, ● represents stations mainly affected by ocean currents |
表1 2022年5月—2023年1月粤东近海浮游动物的种类组成Tab. 1 Species composition of zooplankton in the coastal waters of eastern Guangdong from May 2022 to January 2023 |
| 类群 | 春季种类 | 夏季种类 | 秋季种类 | 冬季种类 | 合计 | 百分比/% |
|---|---|---|---|---|---|---|
| 水螅水母类 Hydrozoa | 8 | 9 | 4 | 3 | 10 | 11.49 |
| 管水母类 Siphonophorae | 3 | 3 | 2 | 1 | 3 | 3.45 |
| 栉水母类 Ctenophores | 1 | 1 | 1 | 0 | 1 | 1.15 |
| 浮游多毛类 Polychaeta | 1 | 1 | 0 | 0 | 1 | 1.15 |
| 浮游软体类 Mollusca | 2 | 2 | 0 | 0 | 2 | 2.30 |
| 枝角类 Cladocera | 1 | 1 | 0 | 0 | 1 | 1.15 |
| 介形类 Ostracoda | 3 | 3 | 1 | 1 | 3 | 3.45 |
| 端足类 Amphipods | 1 | 1 | 1 | 1 | 2 | 2.30 |
| 桡足类Copepoda | 20 | 23 | 12 | 14 | 32 | 36.78 |
| 糠虾类 Mysids | 1 | 1 | 0 | 0 | 1 | 1.15 |
| 磷虾类 Euphausiids | 0 | 0 | 1 | 1 | 2 | 2.30 |
| 十足类 Decapoda | 1 | 2 | 1 | 1 | 2 | 2.30 |
| 毛颚类 Chaetognatha | 7 | 7 | 5 | 3 | 8 | 9.20 |
| 浮游被囊类 Tunicates | 4 | 4 | 0 | 3 | 5 | 5.75 |
| 浮游幼虫 Planktonic larvae | 14 | 14 | 8 | 4 | 14 | 16.09 |
| 合计 Total | 67 | 72 | 36 | 32 | 87 | 100 |
表2 2022年5月—2023年1月粤东近海浮游动物各类群丰度(平均值±标准差, 单位为ind.·m-3)的变化Tab. 2 Variation in the abundance (unit: ind.·m-3) of zooplankton groups in the coastal waters of eastern Guangdong from May 2022 to January 2023 |
| 类群 | 春季丰度 | 夏季丰度 | 秋季丰度 | 冬季丰度 |
|---|---|---|---|---|
| 水母类 Hydrozoa | 3.04±4.14 | 5.22±9.70 | 1.11±1.68 | 0.93±2.07 |
| 桡足类 Copepods | 9.12±12.44 | 36.78±12.44 | 12.72±14.26 | 34.20±40.23 |
| 毛颚类 Chaetognaths | 28.37±31.29 | 17.78±25.23 | 8.37±9.40 | 1.96±3.22 |
| 浮游被囊类 Pelagic tunicates | 8.94±18.39 | 3.51±6.19 | 0 | 2.00±5.29 |
| 浮游幼虫 Planktonic larvae | 41.61±55.11 | 6.54±12.97 | 4.86±5.35 | 5.24±10.41 |
| 其他类群Others | 6.70±11.59 | 7.73±10.63 | 3.53±4.81 | 0.71±0.84 |
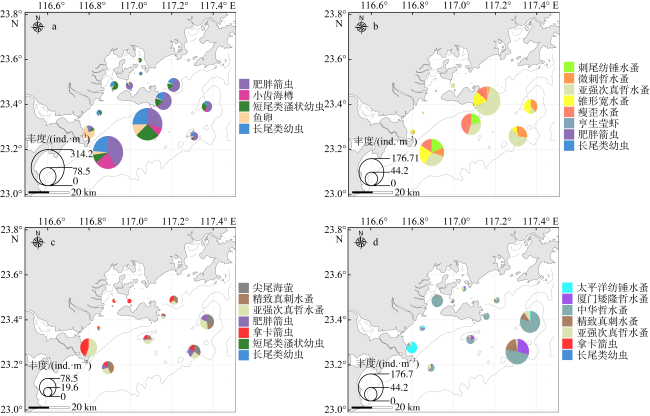
表3 2022年5月—2023年1月粤东近海浮游动物优势种及其丰度(平均值±标准差, 单位为ind.·m-3)的变化Tab. 3 Variation in the abundance (unit: ind.·m-3) of dominant species in the coastal waters of eastern Guangdong from May 2022 to January 2023 |
| 类群 | 优势种 | 春季丰度 | 夏季丰度 | 秋季丰度 | 冬季丰度 |
|---|---|---|---|---|---|
| 介形类 | 尖尾海萤Cypridina acuminata | - | - | 2.80±5.12 | - |
| 桡足类 | 太平洋纺锤水蚤Acartia pacifica | - | - | - | 2.68±7.51 |
| 刺尾纺锤水蚤Acartia spinicauda | - | 2.59±4.81 | - | - | |
| 厦门矮隆哲水蚤Bestiola amoyensis | - | - | - | 3.27 ±8.84 | |
| 中华哲水蚤Calanus sinicus | - | - | - | 18.20±24.38 | |
| 微刺哲水蚤Canthocalanus pauper | - | 3.68±6.20 | - | - | |
| 精致真刺水蚤Euchaeta concinna | - | - | 2.22±3.17 | 3.34±6.76 | |
| 亚强次真哲水蚤Subeucalanus subcrassus | - | 15.20±24.97 | 7.97±12.00 | 2.49±2.59 | |
| 锥形宽水蚤Temora turbinata | - | 5.60±8.58 | - | - | |
| 瘦歪水蚤Tortanus gracilis | - | 4.65±8.08 | - | - | |
| 十足类 | 亨生莹虾Lucifer hanseni | - | 4.67±7.85 | - | - |
| 毛颚类 | 肥胖箭虫Sagitta enflata | 25.07±33.60 | 15.57±21.55 | 1.99±3.14 | - |
| 拿卡箭虫Sagitta nagae | - | - | 5.09±9.22 | 1.32±1.75 | |
| 浮游被囊类 | 小齿海樽Doliolum denticulatum | 7.40±16.79 | - | - | - |
| 浮游幼虫 | 短尾类溞状幼虫Brachyura zoea larvae | 11.91±17.80 | - | 1.64±3.05 | - |
| 鱼卵Fish eggs | 6.94±11.12 | - | - | - | |
| 长尾类幼虫Macrura larvae | 14.19±22.29 | 3.32±7.28 | 2.15±2.70 | 4.32±10.08 |
注: “-”表示在相应航次优势度< 0.02, 未判定为优势种。 |
表4 2022年5月—2023年1月粤东近海不同海区环境因子和浮游动物种数、生物量和丰度的季节变化及检验显著性Tab. 4 Seasonal changes and test significance of environmental factors and zooplankton species number, biomass and abundance in the inshore and offshore waters of eastern Guangdong from May 2022 to January 2023 |
| 季节和显著性检验 | 温度/℃ | 盐度/‰ | 叶绿素a浓度/(μg·L-1) | 浮游动物种数 | 浮游动物生物量 /(mg·m-3) | 浮游动物丰度 /(ind.·m-3) | |
|---|---|---|---|---|---|---|---|
| 近岸 | 春季 | 23.55±0.36 | 28.04±1.69 | 3.22±2.56 | 6±2 | 29.01±21.40 | 17.64±14.14 |
| 夏季 | 27.77±1.88 | 27.55±1.88 | 7.91±3.32 | 3±3 | 10.98±10.80 | 3.33±3.81 | |
| 秋季 | 25.79±0.34 | 29.55±1.40 | 1.81±0.94 | 5±2 | 56.45±63.79 | 18.58±29.80 | |
| 冬季 | 17.11±0.20 | 27.15±2.61 | 3.14±1.10 | 6±1 | 15.58±15.11 | 22.84±19.55 | |
| 远岸 | 春季 | 23.22±0.46 | 31.37±0.71 | 1.88±1.33 | 21±4 | 128.89±94.80 | 178.51±139.49 |
| 夏季 | 26.37±1.66 | 30.95±1.28 | 6.18±5.67 | 23±10 | 340.78±270.80 | 151.80±88.64 | |
| 秋季 | 25.91±0.16 | 31.33±1.47 | 1.07±0.35 | 16±2 | 117.97±115.30 | 42.61±21.70 | |
| 冬季 | 16.88±0.13 | 30.71±0.39 | 2.97±0.36 | 11±4 | 50.02±61.28 | 67.24±73.13 | |
| t-test | P=0.578 | P<0.001 | P=0.112 | P<0.001 | P<0.001 | P<0.001 | |
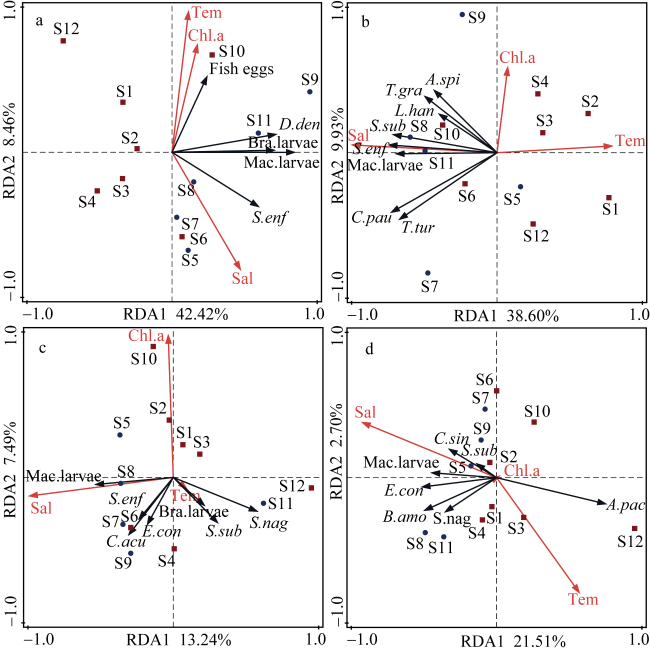
图9 2022年5月—2023年1月粤东近海浮游动物优势种与环境因子的冗余分析a. 春季; b. 夏季; c. 秋季; d. 冬季; 浮游动物优势种缩写的全称请见 Fig. 9 Redundancy analysis of dominant species of zooplankton constrained by environmental factors. (a) spring; (b) summer; (c) autumn; (d) winter; The full name of the abbreviation for the dominant species can be found in the |
| [1] |
蔡德华, 陈振明, 唐书怿, 2020. 南澳岛周边海域海水质量近10年变化趋势浅析[J]. 环境影响评价, 42(2): 63-66.
|
| [2] |
蔡尚湛, 靖春生, 许金电, 等, 2016. 粤东及闽南近岸上升流对局地风场变化的响应[J]. 海洋学报, 38(9): 1-12.
|
| [3] |
陈丹婷, 柯志新, 谭烨辉, 等, 2020. 汕头南澳―东山海域营养盐季节分布特征及其对浮游植物生长的潜在性限制[J]. 生态科学, 39(4): 41-50.
|
| [4] |
杜萍, 徐晓群, 徐旭丹, 等, 2017. 象山港三种不同养殖方式对浮游动物群落的影响[J]. 水产学报, 41(11): 1719-1733.
|
| [5] |
中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员, 2007. GB/T12763. 6-2007海洋调查规范第6部分:海洋生物调查[S]. 北京: 中国标准出版社: 6-37.
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China, 2007. GB 17378. 6-2007 The specification for marine monitoring—Part 6: Marine biological survey[S]. Beijing: Standards Press of China: 6-37 (in Chinese).
|
| [6] |
洪华生, 商少凌, 张彩云, 等, 2005. 台湾海峡生态系统对海洋环境年际变动的响应分析[J]. 海洋学报, 27(2): 63-69.
|
| [7] |
黄长江, 陈善文, 何歆, 等, 2003. 2001—2002年粤东柘林湾浮游动物的生态学研究[J]. 海洋与湖沼, 34(2): 117-130.
|
| [8] |
黄银爽, 欧林坚, 杨宇峰, 2017. 广东南澳岛大型海藻龙须菜与浮游植物对营养盐的竞争利用[J]. 海洋与湖沼, 48(4): 806-813.
|
| [9] |
姜胜, 黄长江, 陈善文, 等, 2002. 2000—2001年柘林湾浮游动物的群落结构及时空分布[J]. 生态学报, 22(6): 828-840.
|
| [10] |
柯志新, 陈丹婷, 谭烨辉, 等, 2019. 汕头南澳-东山海域初级生产力的时空特征[J]. 中国水产科学, 26(1): 44-52.
|
| [11] |
黎素菊, 洪捷娴, 陈树鹏, 2022. 柘林湾养殖区氮、磷季节分布特征及富营养化评价[J]. 江西水产科技, (4): 45-47, 51. (in Chinese)
|
| [12] |
连喜平, 谭烨辉, 黄良民, 等, 2011. 大亚湾大中型浮游动物的时空变化及其影响因素[J]. 海洋环境科学, 30(5): 640-645.
|
| [13] |
刘陈, 魏南, 王庆, 等, 2019. 广东汕头南澳岛近岸海域浮游植物群落结构与环境特征[J]. 应用与环境生物学报, 25(5): 1091-1098.
|
| [14] |
刘萍, 宋洪军, 张学雷, 等, 2015. 桑沟湾浮游动物群落时空分布及养殖活动对其影响[J]. 海洋科学进展, 33(4): 501-511.
|
| [15] |
潘翠红, 夏丽华, 吴志峰, 等, 2021. 柘林湾近岸水产养殖区水域叶绿素a浓度反演[J]. 热带海洋学报, 40(1): 142-153.
|
| [16] |
彭璇, 马胜伟, 陈海刚, 等, 2014. 夏季柘林湾-南澳岛海洋牧场营养盐的空间分布及其评价[J]. 南方水产科学, 10(6): 27-35.
|
| [17] |
任玉正, 柯志新, 谭烨辉, 等, 2020. 广东省南澳岛东部海域浮游动物群落结构及其影响因素[J]. 热带海洋学报, 39(2): 65-76.
|
| [18] |
舒业强, 王强, 俎婷婷, 2018. 南海北部陆架陆坡流系研究进展[J]. 中国科学: 地球科学, 48(3): 276-287.
|
| [19] |
孙鲁峰, 李秀启, 徐兆礼, 2017. 东山湾浮游动物数量特征与养殖活动及水团关系分析[J]. 大连海洋大学学报, 32(4): 465-472.
|
| [20] |
孙松, 李超伦, 张光涛, 等, 2011. 胶州湾浮游动物群落长期变化[J]. 海洋与湖沼, 42(5): 625-631.
|
| [21] |
孙松, 孙晓霞, 2014. 海洋生物功能群变动与生态系统演变[J]. 地球科学进展, 29(7): 854-858.
|
| [22] |
王翠, 郭晓峰, 方婧, 等, 2018. 闽浙沿岸流扩展范围的季节特征及其对典型海湾的影响[J]. 应用海洋学学报, 37(1): 1-8.
|
| [23] |
王亮根, 杜飞雁, 陈丕茂, 等, 2016. 南澳岛北部海域浮游动物生态学特征及水团影响[J]. 南方水产科学, 12(5): 23-33.
|
| [24] |
温浩, 宏波, 2020. 粤东沿岸上升流年际变化及其与渔业相关性分析[J]. 人民珠江, 41(7): 1-11, 17.
|
| [25] |
许金电, 蔡尚湛, 宣莉莉, 等, 2014. 粤东至闽南沿岸海域夏季上升流的调查研究[J]. 热带海洋学报, 33(2): 1-9.
|
| [26] |
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15, 19.
|
| [27] |
义家吉, 颜历, 王洋, 等, 2023. 粤东近岸海域表层沉积物重金属污染评价及来源解析[J]. 海洋环境科学, 42(2): 200-208, 216.
|
| [28] |
曾流明, 1986. 粤东沿岸上升流迹象的初步分析[J]. 热带海洋, 5(1): 68-73.
|
| [29] |
张莹, 2022. 韩江口水体中营养盐的动力过程研究[D]. 汕头: 汕头大学.
|
| [30] |
周枭, 颜秀利, 孙振宇, 等, 2023. 韩江口及邻近海域夏季营养盐和溶解有机质的河口化学特征[J]. 厦门大学学报(自然科学版), 62(3): 385-396.
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}