
Journal of Tropical Oceanography >
Development of testis and ultrastructure of spermatozoa in Mastigias papua
Editor: LIN Qiang
Received date: 2024-01-02
Revised date: 2024-02-08
Online published: 2024-02-28
Supported by
Natural Science Foundation of Xiamen, China(3502Z202372021)
Research Start-up Funds of Jimei University(ZQ2021023)
In this study, Mastigias Papua was selected as the sample, its testis development and spermatozoa ultrastructure was observed in great detail. The development phases of testis and the ultrastructure of the spermatozoa were also studied. The results showed that the nest of M. papus with a bell diameter of (4.99±0.07)cm gradually became matured after 26d-40d under experimental conditions, and the spermatozoa bundle could be continuously discharged for 2-3 months. According to the structural characteristics of spermatozoa follicles and spermatogenesis, the development of the testis can be divided into three stages, namely stage I: the outer part of the testis is not visible to naked eye, the seminal vesicles are small and transparent, and are arranged loosely, and there is no cavity inside; Phase II: the testis is circular, milky white, with gradient development of the gonads. The seminal vesicles that are far from the gastric filament are larger and more tightly arranged, with an internal cavity; Stage III: The folds of the testis are clearly sac-shaped, and under light microscopy the seminal vesicles at different developmental stages can be seen. A large number of seminal vesicles are filled with mature spermatozoa, present in spermbundles. The spermatozoa of M. papus was (50.64±3.93)μm in length, including a sharp chilli shape head (7.61±0.63)μm, a slightly protruding middle section (1.00±0.12)μm. and a very slender tail (43.03±4.02)μm. There was no acrosome in the spermatozoa head, which was mainly composed of nucleus and thin plasma membrane, small vesicles scattered in the plasma membrane. The middle section is composed of two large mitochondria and is connected to the tail by anchoring apparatus. The internal axial filament of the spermatozoa tail was a typical 9+2 duplex microtubule structure.
Key words: Mastigias papua; testis development; spermatozoa ultrastructure
ZHANG Ruixue , YANG Wenhao , CHEN Nan , WANG Shuhong . Development of testis and ultrastructure of spermatozoa in Mastigias papua[J]. Journal of Tropical Oceanography, 2024 , 43(6) : 129 -139 . DOI: 10.11978/2024002
图1 巴布亚硝水母(Mastigias papua)精巢外部形态变化a. 实验开始第10d的精巢; b. 第25d的精巢; c.第40d的精巢; a、b和c中右下角为精巢局部放大; 白色箭头所示为局部放大; 白色三角所示为胃丝; 白色圆点所示为精巢 Fig. 1 Testis morphology changes of Mastigias papua. (a) Testis on the 10th day of the experiment; (b) testis on the 25th day; (c) testis on the 40th day. The lower right corner of (a), (b) and (c) show a partial enlargement of the testis. White arrows show local magnification; white triangles show gastric filaments; white dots show testis |
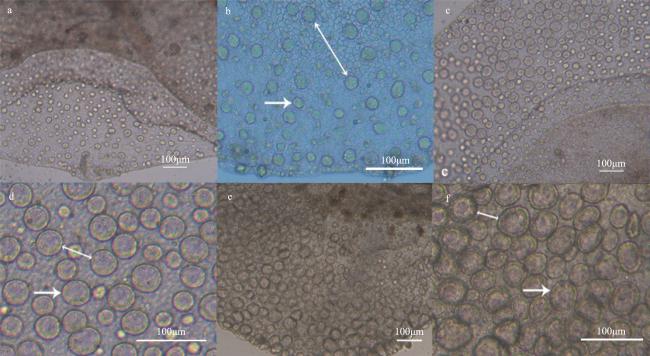
图2 实验第1到10 天巴布亚硝水母(Mastigias papua)的精巢a、c、e分别为培养不同时间巴布亚硝水母的精巢, b、d、f分别为a、c、e局部放大。双箭头表示精小囊间隙; 箭头所示该阶段典型的精小囊 Fig. 2 The testis of Mastigias papua from 1 to 10d of the experiment. (a, c, e) The testis of Mastigias papua sampled from different periods, respectively; (b), (d) and (f) are local enlargements of (a), (c) and (e), respectively. The double arrows show the gap of spermatozoa follicles, the arrow shows typical spermatozoa follicles for this phase |
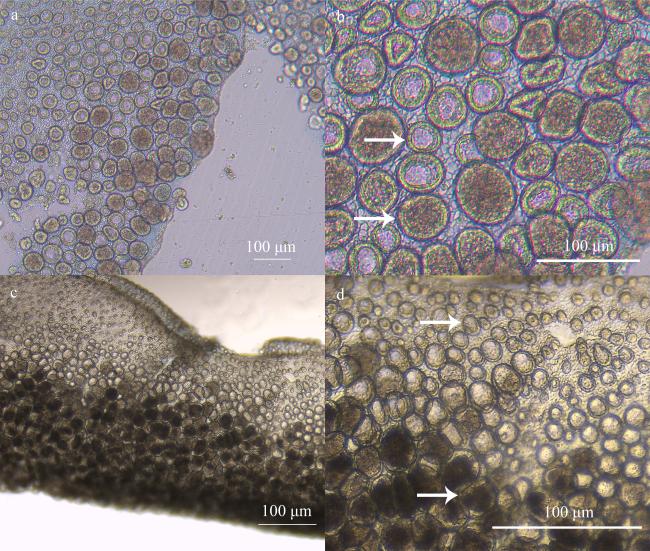
图3 实验第11到25 天巴布亚硝水母(Mastigias papua)的精巢a和c分别为培养不同时间巴布亚硝水母的精巢, b和d分别为a和c放大的精小囊。箭头所示该阶段典型的精小囊 Fig. 3 The testis of Mastigias papua from 11 to 25d of the experiment. (a, c) The testis of Mastigias papua sampled from different period, respectively; (b) and (d) are the enlarged spermatozoa follicles of (a) and (c), respectively. The arrows show typical spermatozoa follicles for this phase |
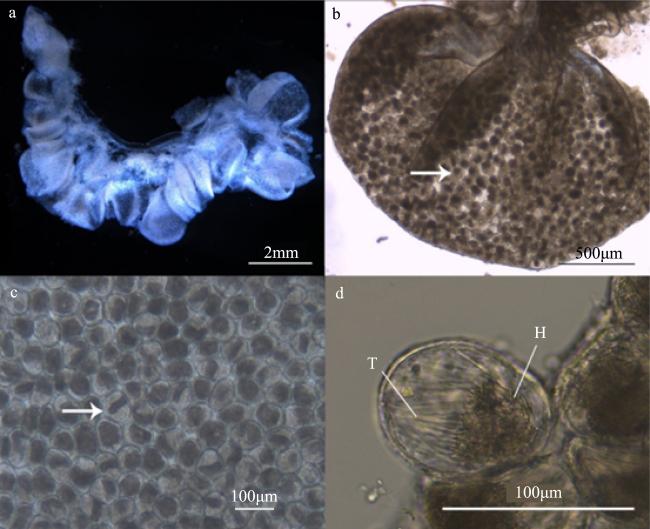
图4 实验第26到40天巴布亚硝水母(Mastigias papua)的精巢a. 饱满的性腺 b. 囊状精巢 c. 不透明的精小囊 d. 精小囊中精子束。H: 精子头部; T: 精子尾部; 箭头所示处为该阶段性腺典型精小囊 Fig. 4 The testis of Mastigias papua from 26 to 40d of the experiment. (a) Full gonads; (b) the shape of a sac testis; (c) the opaque spermatozoa follicles; (d) spermatozeugmatas in the spermatozoa follicle. H: head of spermatozoa; T: tail of spermatozoa. The arrows show the typical spermatozoa follicles for this phase |
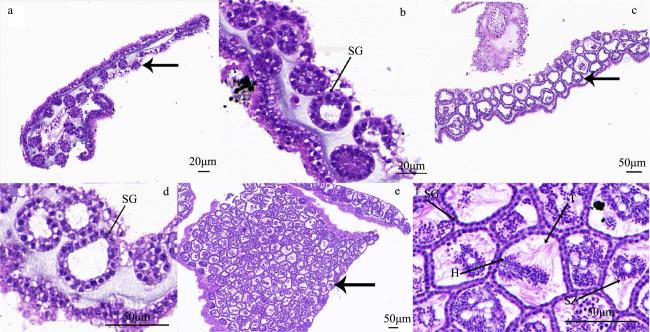
图5 巴布亚硝水母(Mastigias papua)不同阶段精巢组织结构a、b: 实验开始第1—10d的精巢; c、d: 第11—25d的精巢; e、f: 第26—40d的精巢; H: 精子头部; SG: 精原细胞; SZ: 精子; T: 精子尾部; 粗箭头所示为精巢不同发育时期的精小囊 Fig. 5 Histology of various stages of Mastigias papua testis. (a, b) The testis from 1 to 10d of the experiment; (c, d) the testis from 11 to 25d of the experiment; (e, f) the testis from 26 to 40d of the experiment. H: head of spermatozoa; SG: spermatogonia; SZ: spermatozoa; T: tail of spermatozoa. The arrows show the spermatozoa follicles at different stages of testis development |
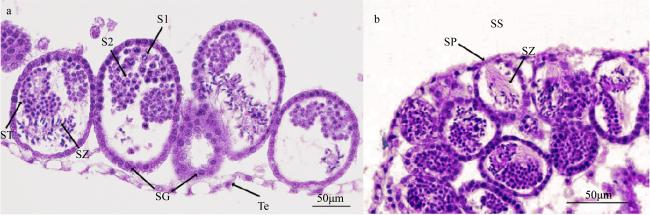
图6 巴布亚硝水母(Mastigias papua)不同发育时期精小囊的组织结构a. 发育早期的精小囊; b. 发育晚期的精小囊。SG: 精原细胞; S1: 初级精母细胞; S2: 次级精母细胞; SS: 生殖下穴; SP: 排精坑; ST: 精细胞; SZ: 精子; Te: 生殖上皮 Fig. 6 Spermatozoa follicles structure from testis at different developmental stages (Mastigias papua). (a) Early development of spermatozoa follicles; (b) late development of spermatozoa follicles. SG: spermatogonia; S1: primary spermatocytes; S2: secondary spermatocytes; SZ: spermatozoa; SS: subgenital sinus; SP: spermiation pit; ST: spermatids; Te: testicular epithelium |
图7 巴布亚硝水母(Mastigias papua)精子形态a. 成熟精子完整结构; b、c. 精子头部和中段。Aa: 锚定装置; H: 头部; M: 线粒体; Ms: 中段; T: 尾部 Fig. 7 Mastigias papua spermatozoas morphology. (a) Complete structure of mature spermatozoa; (b, c) the head and mid-section of mature spermatozoa. Aa: anchoring apparatus; H: head; M: mitochondria; Ms: middle section; T: tail |
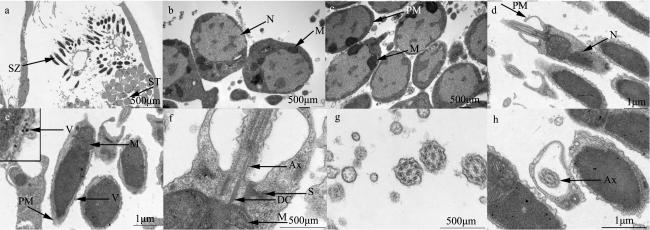
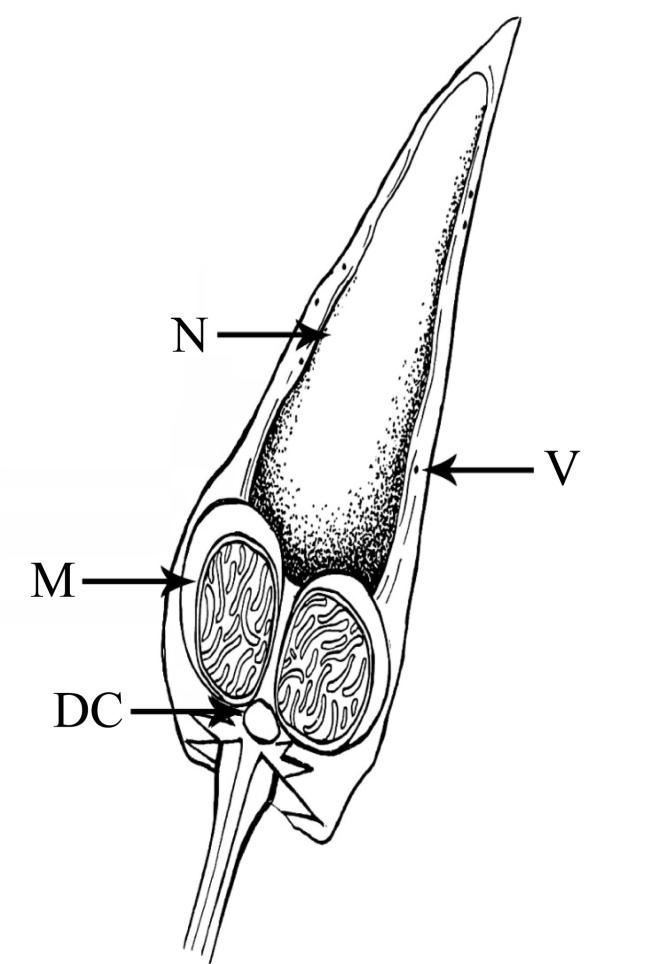
图8 巴布亚硝水母(Mastigias papua)精子的超微结构a. 性腺中精细胞和成熟精子; b. 精细胞; c. 成熟精子; d—f. 成熟精子头部和中段; g、h. 成熟精子尾部横切图。Ax: 轴丝; DC: 远端中心粒; S: 卫星状微丝; ST: 精细胞; SZ: 精子; PC: 近端中心粒; PM: 质膜; M: 线粒体; Mi: 微管; N: 细胞核; V: 囊泡 Fig. 8 Untrostructure of spermatozoas from Mastigias papua. (a) Spermatids and mature spermatozoas in the gonad; (b) shows spermatids; (c) shows mature spermatozoas; (d-f) the head and mid-section of mature spermatozoas; (g, h) cross-sections of the tails of mature spermatozoas. Ax: axoneme; DC: distal centriole; S: satellite rays; ST: spermatids; SZ: spermatozoa; PC: proximal centriole; PM: plasma membrane; M: mitochondria; Mi: microtubule; N: nucleus; V: vesicles |
表1 巴布亚硝水母(Mastigias papua)精巢不同时期发育情况Tab. 1 Development of the testis of Mastigias papua at different stages |
| 精巢分期 | 外部形态 | 光学显微镜下 | 组织切片 |
|---|---|---|---|
| Ⅰ期 | 肉眼不可见(图1a) | 精小囊小而透明, 排列松散(图2) | 由数个细胞核明显的精原细胞组成的球状精小囊, 内部无空腔(图5a、b) |
| Ⅱ期 | 环状乳白色, 由近胃丝处向外性腺透明度降低、颜色加深(图1b) | 梯度发育, 由近胃丝处向外精小囊个体变大, 透明度降低(图3) | 远离胃丝的精小囊更大, 排列更紧密, 内部有空腔, 可见明显的由精原细胞整齐排列而成的精小囊壁(图5c、d) |
| Ⅲ期 | 褶皱呈囊状, 浅米黄色(图1c) | 处于不同发育阶段的精小囊, 正在进行精子发生的精小囊占绝大多数(图4) | 精小囊内有不同发育阶段的配子细胞, 大量精小囊内充满以精子束存在的成熟精子(图5e、f) |
表2 部分钵水母纲精子外部形态Tab. 2 Scyphozoa spermatozoa morphology |
| [1] |
陈昭廷, 李琪, 陈四清, 等, 2015. 海月水母精巢发育及排精过程的观察[J]. 动物学杂志, 50(1): 131-140.
|
| [2] |
陈昭廷, 周洋, 顾志峰, 等, 2017. 海月水母(Aurelia aurita)精巢发育及精子的超微结构[J]. 海洋与湖沼, 48(1): 122-129.
|
| [3] |
洪惠馨, 2014. 中国海域钵水母生物学及其与人类的关系[M]. 北京: 海洋出版社:60.
|
| [4] |
杨翠华, 王玮, 王文章, 2011. 光照和水流对巴布亚硝水母存活时间及存活率的影响[J]. 水产科学, 30(9): 555-558.
|
| [5] |
杨翠华, 王玮, 王文章, 等, 2012. 咖啡金黄水母的人工培育及生活史观察[J]. 水产科学, 31(12): 708-712.
|
| [6] |
杨文浩, 王淑红, 林子路, 等, 2022. 巴布亚硝水母无性生殖过程及形态变化[J]. 动物学杂志, 57(3): 321-335.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}