
Journal of Tropical Oceanography >
Phytoplankton community structure and its relationship with environmental factors in the spring coastal region of Nan’ao based on morphology and high-throughput sequencing
Copy editor: SUN Cuici
Received date: 2024-02-28
Revised date: 2024-03-25
Online published: 2024-05-17
Supported by
Special Fund for Science and Technology Planning Project of Guangdong Province of China(2021B1212050023)
Guangdong Basic and Applied Basic Research Foundation(2022A1515010656)
Guangzhou Municipal Key Science and Technology Project(2023B03J1328)
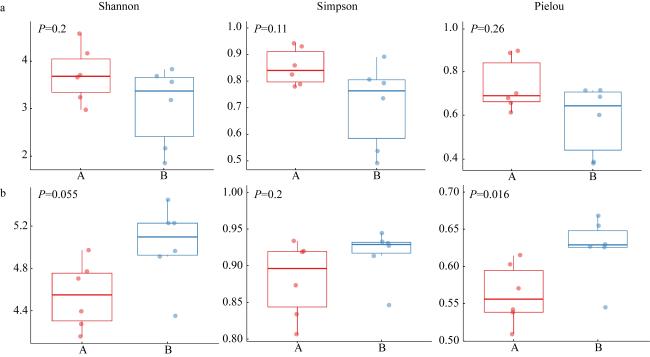
The structure of phytoplankton community was investigated based on morphological identification and high-throughput sequencing in the Nan’ao waters in May 2022. The spatial distribution of phytoplankton in relation to environmental factors was explored and results from two different methods were also compared and discussed. According to the cluster analysis of environmental factors, the survey area can be divided into nearshore area and offshore area. Totally, 105 species belong to 52 genera, 39 families, 27 orders, 8 orders, 5 phyla were morphological identified, while 543 species in 6 phyla, 32 orders, 97 families, 155 families, 272 genera through high-throughput sequencing were identified. Bacillariophyta and Dinophyta were the main groups, composing 74.54% and 24.78% through microscopic identification, and 17.52% and 67.84% through high-throughput sequencing, respectively. The abundance of phytoplankton identified by morphology ranged from 0.11×105 to 6.85×105 cells·L-1, which was significantly correlated with the distribution of chlorophyll a concentration, and the phytoplankton diversity index was lower than that of the high-throughput sequencing method. 7 dominant species were obtained by both methods, of which the absolutely dominant specie in the morphological identification results was Chaetoceros compressus, and the absolute dominant specie in the high-throughput sequencing identification results was Heterocapsa rotundata. Correlation analysis showed that the main environmental factor affecting phytoplankton community structure in Nan’ao in spring was pH, salinity and DIP (dissolved inorganic phosphorus). Using the combination of morphological identification and high-throughput sequencing technology, the structure and diversity of the phytoplankton community can be described more comprehensively and accurately.
ZHOU Zhixi , TANG Huijuan , KE Zhixin , LIU Jiaxing , ZHOU Weihua . Phytoplankton community structure and its relationship with environmental factors in the spring coastal region of Nan’ao based on morphology and high-throughput sequencing[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 53 -65 . DOI: 10.11978/2024046
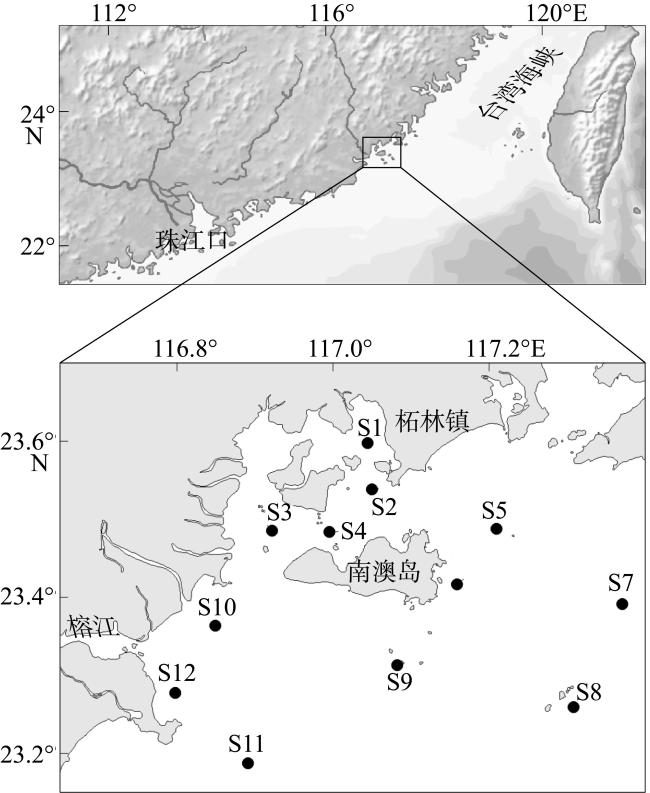
图2 表层环境因子聚类的调查站位分布(a)和聚类结果(b) (●为近岸组, ■为远岸组, 审图号同图1)Fig. 2 Distribution of sampling stations based on surface environmental factors clustering (a, ● represents stations of nearshore group, ■ represents stations of offshore group) and clustering results (b) |
表1 2022年5月南澳近海不同海域环境因子差异及检验显著性Tab. 1 Variability and test significance of environmental factors in the nearshore and offshore waters of Nan’ao in May 2022 |
| 因子 | 温度/℃ | 盐度/‰ | pH | 浊度/NTU | DIN/(μmol·L-1) | DIP/(μmol·L-1) | DSi/(μmol·L-1) | 氮磷比 |
|---|---|---|---|---|---|---|---|---|
| 近岸 | 23.55±0.39 | 28.04±1.85 | 8.03±0.09 | 4.17±2.63 | 12.31±4.39 | 0.77±0.59 | 24.56±4.55 | 26.13±21.53 |
| 远岸 | 23.22±0.50 | 31.37±0.78 | 8.16±0.06 | 1.93±2.22 | 2.06±0.64 | 0.16±0.09 | 7.20±3.61 | 105.08±18.47 |
| t-test | t=1.528, P=0.187 | t=5.126, P<0.01 | t=5.665, P<0.01 | t=1.53, P=0.187 | t=5.81, P<0.05 | t=2.95, P<0.05 | t=8.93, P<0.01 | t=17.45, P<0.01 |
表2 2022年5月南澳海域浮游植物优势种Tab. 2 Dominant species in the coastal waters of Nan’ao in May 2022 |
| 分类水平 | 优势度 | 出现频率% | ||
|---|---|---|---|---|
| 门 | 种 | 形态学鉴定 | 高通量测序 | |
| 硅藻门Bacillariophyta | 扁面角毛藻Chaetoceros compressus | 0.33 | - | 83.33 |
| 微眼藻Minutocellus sp. | - | 0.05 | 100 | |
| 多形微眼藻Minutocellus polymorphus | - | 0.03 | 100 | |
| 艾氏角毛藻Chaetoceros eibenii | 0.02 | - | 58.33 | |
| 洛氏菱形藻Nitzschia lorenziana | 0.02 | - | 83.33 | |
| 尖刺拟菱形藻Pseudo-nitzschia pungens | 0.02 | - | 100 | |
| 中肋骨条藻Skeletonema costatum | 0.02 | - | 41.67 | |
| 甲藻门 Dinophyta | 球形异帽藻Heterocapsa rotundata | - | 0.22 | 100 |
| 海洋原甲藻Prorocentrum micans | 0.13 | - | 100 | |
| 剧毒卡洛藻Karlodinium veneficum | - | 0.09 | 100 | |
| 单眼藻Warnowia sp. | - | 0.09 | 100 | |
| 单眼藻Warnowia spp. | 0.07 | 100 | ||
| 锥状斯氏藻Scrippsiella trochoidea | 0.02 | - | 83.33 | |
| 绿藻门Chlorophyta | 共球藻目Picochlorum sp. | - | 0.03 | 100 |
注: “-”表示在相应航次优势度< 0.02, 未判定为优势种 |
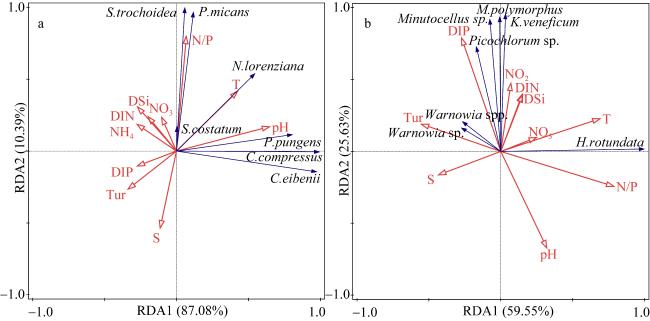
图6 形态学鉴定(a)和高通量测序(b)的浮游植物优势种相对丰度及分布情况C. compressus为扁面角毛藻, C. eibenii为艾氏角毛藻, N. lorenziana为洛氏菱形藻, P. pungens为尖刺拟菱形藻, S. costatum为中肋骨条藻, P. micans为海洋原甲藻, S. trochoidea锥状斯氏藻, H. rotundata为球形异帽藻, K. veneficum为剧毒卡洛藻, Warnowia sp.和Warnowia spp.为单眼藻, Minutocellus sp. 为微眼藻, Picochlorum sp. 属于共球藻目, M. polymorphus为多形微眼藻 Fig. 6 Relative abundances and distributions of dominant species by morphological (a) and molecular (b) analyses |
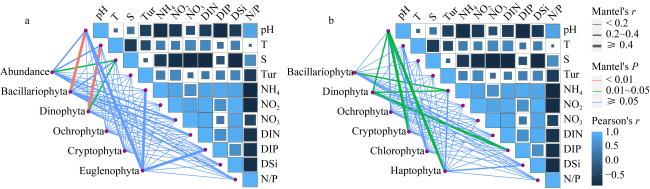
图7 基于形态学鉴定(a)和高通量测序(b)浮游植物群落门水平上与环境因子的相关性Fig. 7 Pairwise comparison of the phytoplankton community and environmental factors based on morphological (a) and molecular (b) analyses |
| [1] |
蔡德华, 陈振明, 唐书怿, 2020. 南澳岛周边海域海水质量近10年变化趋势浅析[J]. 环境影响评价, 42(2): 63-66.
|
| [2] |
陈丹婷, 柯志新, 谭烨辉, 等, 2020. 汕头南澳-东山海域营养盐季节分布特征及其对浮游植物生长的潜在性限制[J]. 生态科学, 39(4): 41-50.
|
| [3] |
陈菊芳, 齐雨藻, 徐宁, 等, 2005. 大亚湾拟菱形藻水华及其在生物群落中的生态地位[J]. 海洋学报, 27(1): 114-119.
|
| [4] |
陈炜婷, 姜广甲, 邓伟, 等, 2023. 基于GIS的2001-2020年广东近岸海域赤潮时空分布[J]. 环境工程, 41(S2): 49-53.
|
| [5] |
杜虹, 王亮根, 曹会彬, 等, 2011. 汕头港浮游植物组成特征及其与环境的关系[J]. 生态学杂志, 30(8): 1757-1765.
|
| [6] |
郭玉洁, 2003. 中国海藻志[M]. 北京: 科学出版社 (in Chinese).
|
| [7] |
黄长江, 董巧香, 郑磊, 1999. 1997年底中国东南沿海大规模赤潮原因生物的形态分类与生态学特征[J]. 海洋与湖沼, 30(6): 581-590.
|
| [8] |
黄圆, 岑竞仪, 梁芊艳, 等, 2024. 基于高通量测序技术的深圳湾真核浮游植物群落结构研究[J]. 热带海洋学报, 43(2): 21-33.
|
| [9] |
暨卫东, 黄尚高, 1990. 台湾海峡西部营养盐变化特征-Ⅲ. 水系混合及浮游植物摄取转移对磷酸盐含量变化影响的统计分析[J]. 海洋学报, 12(4): 447-454 (in Chinese).
|
| [10] |
雷光英, 杨宇峰, 李宵, 2010. 龙须菜对赤潮异弯藻和海洋原甲藻的生长抑制效应[J]. 海洋环境科学, 29(1): 27-31.
|
| [11] |
李冀刚, 杨振雄, 葛颂, 等, 2023. 汕头韩江榕江河口近岸海域浮游植物群落结构与环境因子的关系[J]. 海洋湖沼通报, 45(2): 133-141.
|
| [12] |
黎素菊, 洪捷娴, 陈树鹏, 2022. 柘林湾养殖区氮、磷季节分布特征及富营养化评价[J]. 江西水产科技, (4): 45-47, 51 (in Chinese).
|
| [13] |
李欣, 孙军, 田伟, 等, 2012. 2009年夏季南海北部的浮游植物群落[J]. 海洋科学, 36(10): 33-39.
|
| [14] |
林更铭, 杨清良, 2021. 西太平洋浮游植物物种多样性[M]. 北京: 科学出版社.
|
| [15] |
刘陈, 魏南, 王庆, 等, 2019. 广东汕头南澳岛近岸海域浮游植物群落结构与环境特征[J]. 应用与环境生物学报, 25(5): 1091-1098.
|
| [16] |
刘卫东, 宋伦, 吴景, 2017. 环境样本中微型和微微型浮游植物高通量测序的引物优化[J]. 生态学报, 37(12): 4208-4216.
|
| [17] |
刘旭平, 单忠才, 李媛芳, 等, 2023. 滤食性贝类选择性摄食行为研究现状[J]. 特种经济动植物, 26(5): 89-92, 107 (in Chinese).
|
| [18] |
马华栋, 王平, 温玉波, 等, 2021. 基于生态足迹分析的南澳岛旅游业可持续发展评估[J]. 海洋开发与管理, 38(8): 61-66.
|
| [19] |
彭璇, 马胜伟, 陈海刚, 等, 2014. 夏季柘林湾-南澳岛海洋牧场营养盐的空间分布及其评价[J]. 南方水产科学, 10(6): 27-35.
|
| [20] |
宋荪阳, 黄金臣, 岳强, 等, 2023. 唐山市海水养殖尾水监测及分析[J]. 河北渔业, (7): 23-29.
|
| [21] |
苏纪兰, 王卫, 1990. 南海域台湾暖流源地问题[J]. 东海海洋, 8(3): 1-9.
|
| [22] |
孙军, 刘东艳, 2004. 多样性指数在海洋浮游植物研究中的应用[J]. 海洋学报, 26(1): 62-75.
|
| [23] |
孙军, 2007. 今生颗石藻的有机碳泵和碳酸盐反向泵[J]. 地球科学进展, 22(12): 1231-1239.
|
| [24] |
谭香, 夏小玲, 程晓莉, 等, 2011. 丹江口水库浮游植物群落时空动态及其多样性指数[J]. 环境科学, 32(10): 2875-2882.
|
| [25] |
唐涛, 蔡庆华, 刘建康, 2002. 河流生态系统健康及其评价[J]. 应用生态学报, 13(9): 1191-1194.
|
| [26] |
王艳, 聂瑞, 李扬, 等, 2010. 广东沿海角毛藻(Chaetoceros)的种类多样性及其地理分布[J]. 海洋科学进展, 28(3): 342-352.
|
| [27] |
王雨, 林茂, 林更铭, 等, 2011. 闽粤近岸夏季上升流区浮游植物群落组成及其影响因素[J]. 应用生态学报, 22(2): 503-512.
|
| [28] |
吴转璋, 朱超, 唐萍, 等, 2023. 巢湖湖区浮游植物群落与水质因子相关性分析[J]. 生物学杂志, 40(1): 79-84.
|
| [29] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员, 2007. GB/T12763. 6-2007海洋调查规范第6部分:海洋生物调查 [S]. 北京: 中国标准出版社: 6-37.
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China, 2007. GB/T12763. 6-2007 Specifications for oceanographic survey—Part 6: Marine biological survey[S]. Beijing: Standards Press of China: 6-37 (in Chinese).
|
| [30] |
中华人民共和国生态环境部,2020. 近岸海域环境监测规范(HJ 442. 3-2020)第三部分近岸海域水质监测[S]. 北京: 中国环境出版集团出版.
Ministry of Ecology and Environment of the People’s Republic of China, 2020. Technical specification for offshore environmental monitoring Part 3 offshore seawater quality monitoring[S]. Beijing: China Environmental Science Press.
|
| [31] |
周凯, 黄长江, 姜胜, 等, 2002. 2000-2001年柘林湾浮游植物群落结构及数量变动的周年调查[J]. 生态学报, 22(5): 688-698.
|
| [32] |
朱小山, 吴玲玲, 杨瑶, 等, 2005. 粤东柘林湾增养殖区氮磷的分布特征及其富营养化状态评价[J]. 海洋湖沼通报, (3): 16-22.
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}