
Journal of Tropical Oceanography >
MaxEnt model predicting potential distribution of Trichiurus japonicus in the coastal waters of China under global climate change
Received date: 2024-12-28
Revised date: 2025-03-18
Online published: 2025-03-19
Supported by
Sustainable Development Program of Shenzhen(2023N066)
National Natural Science Foundation of China(41976108)
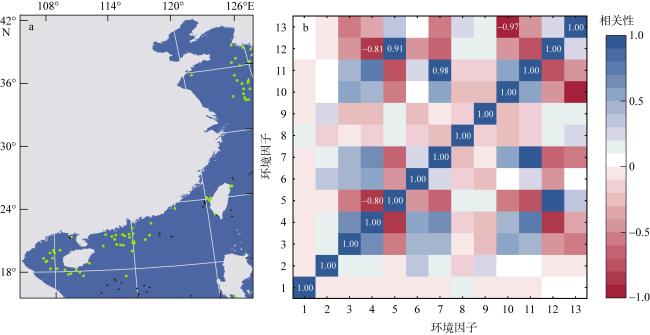
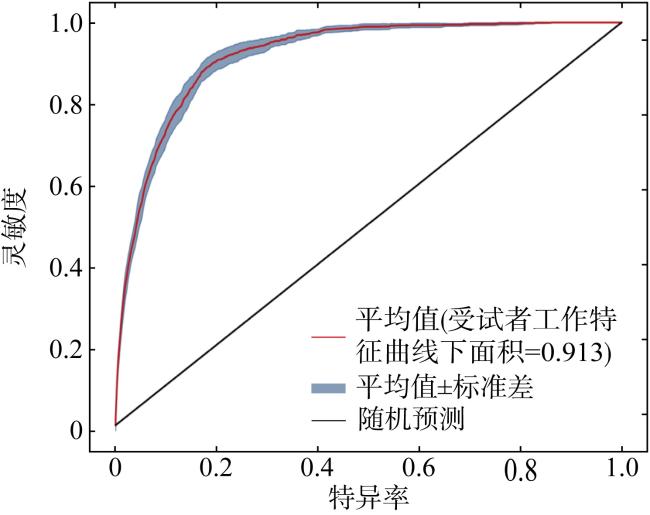
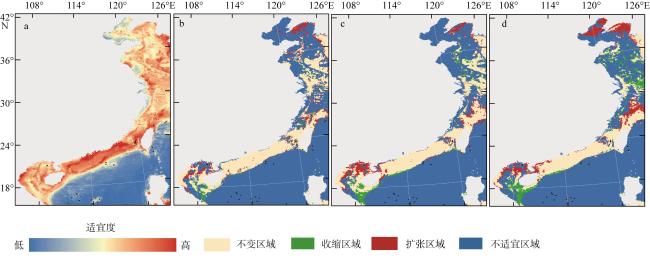
This study predicted the potential distribution of Trichiurus japonicus along China’s coastal waters under the influence of global climate change, using the maximum entropy (MaxEnt) model integrated with Geographic Information System (GIS) techniques. Species occurrence data (70 valid points) were obtained from Global Biodiversity Information Facility (GBIF) and FishBase, while environmental variables were sourced from Bio-ORACLE. Model performance was evaluated using the Receiver Operating Characteristic (ROC) curve, yielding a high accuracy (0.913) of Area Under the Curve (AUC). Our results indicated that suitable habitats for T. japonicus are distributed across China’s four major marine regions, with medium-to-high suitability areas accounting for 11.96% of the total predicted area. Temperature, chlorophyll concentration, and primary productivity were identified as key environmental factors affecting hairtail distribution. Model projections under different shared socioeconomic pathway (SSP) scenarios suggested an expansion of suitable habitats with a potential northward shift towards the Yellow Sea and Bohai Sea, while contracting in the waters of South China such as Beibu Gulf in the future.
FENG Zhanquan , SU Maoliang , DU Yuanyuan , ZHONG Youling , ZHANG Junbin . MaxEnt model predicting potential distribution of Trichiurus japonicus in the coastal waters of China under global climate change[J]. Journal of Tropical Oceanography, 2025 , 44(5) : 77 -85 . DOI: 10.11978/2024241
表1 来自Bio-ORACLE的环境变量Tab. 1 Environmental variables from Bio-ORACLE |
| 因子序号 | 环境因子 | 单位 |
|---|---|---|
| bio_1 | 底层海流方向bot current direction | ° |
| bio_2 | 底层海流速度bot current velocity | m·s-1 |
| bio_3 | 底层溶解氧bot dissolved oxygen | mmol·m-3 |
| bio_4 | 底层初级生产力bot primary productivity | mmol·m-3 |
| bio_5 | 底层盐度bot salinity | ‰ |
| bio_6 | 底层温度bot temperature | ℃ |
| bio_7 | 表层叶绿素surf chlorophyll | mg·m-3 |
| bio_8 | 表层海流方向surf current direction | ° |
| bio_9 | 表层海流速度surf current velocity | m·s-1 |
| bio_10 | 表层溶解氧surf dissolved oxygen | mmol·m-3 |
| bio_11 | 表层初级生产力surf primary productivity | mmol·m-3 |
| bio_12 | 表层盐度surf salinity | ‰ |
| bio_13 | 表层温度surf temperature | ℃ |
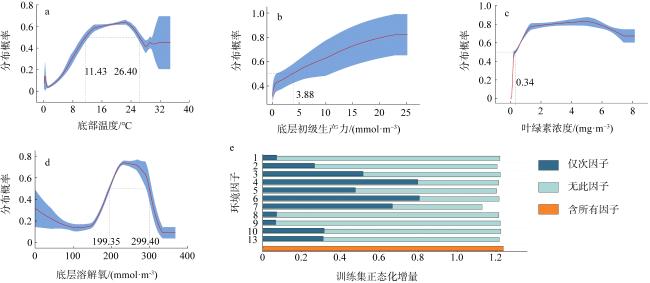
图3 底层温度响应曲线(a)、底层初级生产力响应曲线(b)、叶绿素浓度响应曲线(c)、底层溶解氧响应曲线(d)和基于刀切法的环境因子重要性检验(e)图a—d中的数字为分布概率在0.50之上的环境因子临界值, 红色曲线为平均值, 蓝色区域为平均值±标准差 Fig. 3 Response curve of bottom temperature (a); response curve of bottom primary productivity (b); response curve of chlorophyll concentration (c); response curve of bottom dissolved oxygen (d); environmental factor importance validation using Jackknife Test (e) |
图4 基于不同SSP情景的中国近海带鱼适生区分布预测a. 当前带鱼适生区分布; b、c、d分别表示在SSP1-1.9、SSP2-4.5和SSP5-8.5情景下的未来适生区分布变化预测。该图基于自然资源部标准地图服务网站下载的审图号为GS (2016) 1550号的标准地图制作 Fig. 4 Predicted suitable habitat distribution of T. japonicus in coastal waters of China under different SSPs. (a) Current distribution of suitable habitats for T. japonicus; (b), (c), and (d) represent predicted changes in future suitable habitat distribution under the SSP1-1.9, SSP2-4.5, and SSP5-8.5, respectively |
表2 中国近海带鱼适生区分布概率分级表Tab. 2 Classification of distribution probability grades for T. japonicus suitable habitats in coastal waters of China |
| 区域 | 现在/% | SSP1-1.9/% | SSP2-4.5/% | SSP5-8.5/% |
|---|---|---|---|---|
| 不适宜区(0~MPT) | 16.03 | 19.58 | 22.17 | 10.79 |
| 边缘适生区(MPT~10P) | 52.10 | 48.00 | 45.85 | 56.07 |
| 低适生区(10P~0.5) | 19.90 | 18.99 | 18.25 | 19.63 |
| 中适生区(0.5~0.75) | 11.47 | 12.76 | 12.98 | 12.22 |
| 高适生区(0.75~1) | 0.49 | 0.68 | 0.75 | 1.29 |
| [1] |
曹睿星, 官文江, 高峰, 等, 2023. 基于最大熵和栖息地指数模型预测东、黄海日本鲭渔场分布[J]. 海洋学报, 45(9): 72-81.
|
| [2] |
杜萍, 陈全震, 李尚鲁, 等, 2020. 东海带鱼资源变动及其栖息地驱动因子研究进展[J]. 广东海洋大学学报, 40(1): 126-132.
|
| [3] |
刘星雨, 2022. 气候变化对中国近海主要经济鱼类潜在适宜生境影响的研究[D]. 舟山: 浙江海洋大学.
|
| [4] |
马腾龙, 万亿, 吴萍娟, 等, 2024. 环境因素对浮游植物生长及群落结构的影响[J]. 环境保护前沿, (3): 459-466.
|
| [5] |
齐国君, 陈婷, 高燕, 等, 2015. 基于Maxent的大洋臀纹粉蚧和南洋臀纹粉蚧在中国的适生区分析[J]. 环境昆虫学报, 37(2): 219-223.
|
| [6] |
孙述好, 黄紫薇, 胡男, 等, 2024. 大麻哈鱼幼鱼对温度、盐度、pH及溶解氧耐受性的研究[J]. 黑龙江水产, 43(2): 140-143.
|
| [7] |
武晓宇, 董世魁, 刘世梁, 等, 2018. 基于MaxEnt模型的三江源区草地濒危保护植物热点区识别[J]. 生物多样性, 26(2): 138-148.
|
| [8] |
张曼, 2022. 北部湾带鱼资源状况研究[D]. 上海: 上海海洋大学.
|
| [9] |
张曼, 王雪辉, 蔡研聪, 等, 2022. 北部湾带鱼空间聚散变化特征[J]. 中国水产科学, 29(11): 1647-1658.
|
| [10] |
张鹏, 2020. 阿曼湾海域带鱼生物学特性、矢耳石形态和微化学研究[D]. 上海: 上海海洋大学.
|
| [11] |
张旭, 张佳, 王婕, 等, 2023. 温度和光照周期对硬头鳟幼鱼生长、生理及行为的影响[J]. 大连海洋大学学报, 38(2): 251-258.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}