
Journal of Tropical Oceanography >
Skeletal microstructure observations and calcification process analysis of two species of Montipora
Received date: 2016-02-15
Request revised date: 2016-09-08
Online published: 2017-01-19
Supported by
National Natural Science Foundation of China (31460555)
State Oceanic Administration Project (HZ2012-174)
“Central and Western Colleges and Universities Enhance the Comprehensive Strength” Funding Projects
Copyright
Montipora is the second species-rich scleractinian genus that is widely distributed in the Indo-Pacific Ocean. Montipora digitata and Montipora fragilis, two species of branching Montipora, from the South China Sea were selected in this study, and its skeletal microstructures and calcification processes were observed and analyzed using Scanning Electron Microscopy (SEM) combined with Energy Dispersive Spectrometer (EDS). The results showed that the two species of Montipora skeletal basic elements were substantially the same (including calice, septum, theca, dissepiment, etc.), but there were many differences in details. Coenosteum surface of M. digitata was glabrous and recticular formation with shorter spines (about 40 μm) and each calices (about 1 mm) were separated by big interval. The septa of the first cycle was poorly developed except that the shape of its direct septum was sheet. In addition, teeth along the margin of septa developed well and had flat shape. The coenosteum surface of M. fragilis was tuberculate and recticular formation with many small spines (about 100 μm) and irregular nodular ridge between the calices. Compared with M. digitata, the first-cycle septa of M. fragilis had degenerated teeth. Main ingredients of the two kinds of coral skeletons were aragonite crystals, with most being tufted-crystals and a few being pebble-crystals. EDS analysis showed that the components of calcium carbonate from the skeleton were not directly generated, but formed in the following four processes: 1) the calcium was concentrated to develop calcium point; 2) calcium combined with carbon and oxygen, and the content of carbon was higher than that of oxygen; 3) aragonite crystals tended to look regular, and the content of carbon decreased and that of oxygen increased; and 4) a lot of aragonite crystals gathered into sclerodermite in orderly arrangement to form mature skeleton.
GENG Taonian , YAO Xuemei , ZHANG Ying , XIE Xialing , CUI Min , LIN Daoming . Skeletal microstructure observations and calcification process analysis of two species of Montipora[J]. Journal of Tropical Oceanography, 2017 , 36(1) : 56 -64 . DOI: 10.11978/2016015
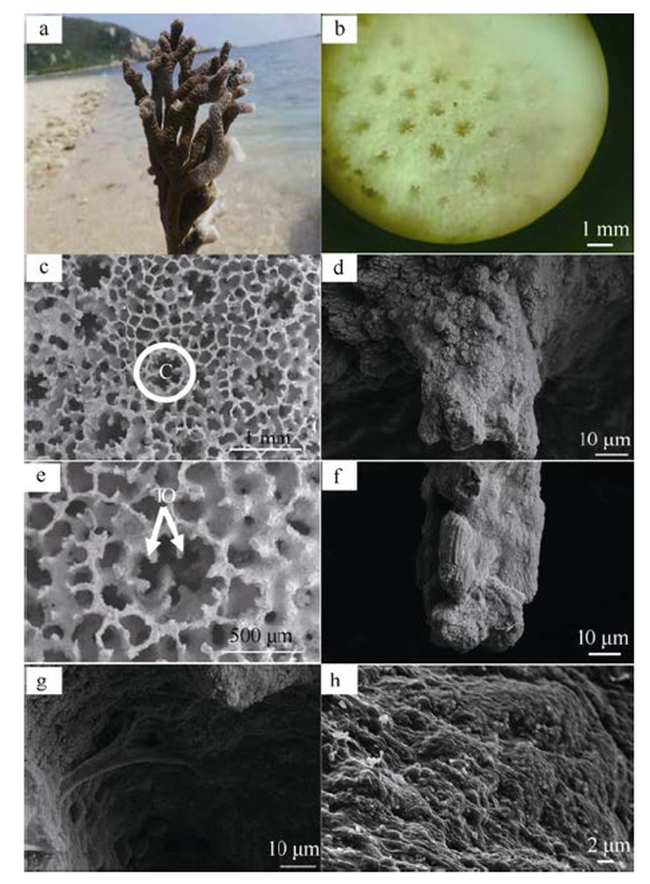
Fig. 1 Skeletal basic structure unit of M. digitata图1 指状蔷薇珊瑚骨骼基本结构单元 |
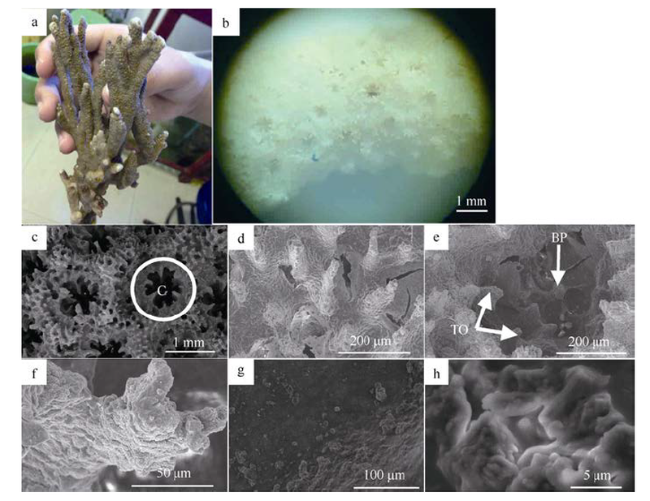
Fig. 2 Skeletal basic structure unit of M. fragilis图2 脆蔷薇珊瑚骨骼基本结构单元 |
Fig. 3 Skeletal shadow branch surface structures of Montipora digitata and Montipora fragilis图3 指状蔷薇珊瑚与脆蔷薇珊瑚骨骼背阴面表面结构 |
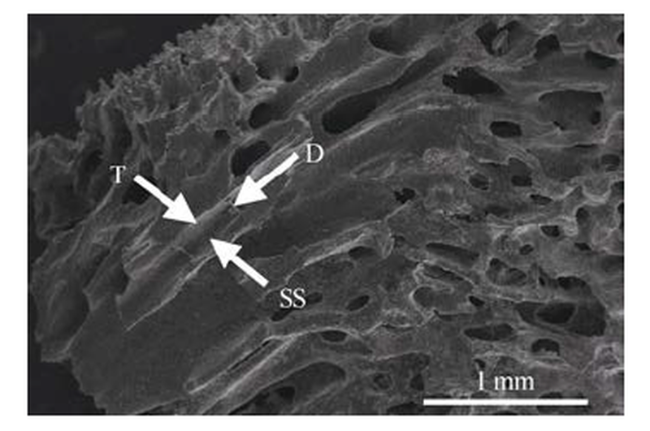
Fig. 4 Skeletal internal structure characteristics of M. digitata and M. fragilis图4 指状蔷薇珊瑚与脆蔷薇珊瑚骨骼内部结构特征 |
Fig. 5 Microstructure of M. fragilis growing point图5 脆蔷薇珊瑚的生长点表观结构 |
Fig. 6 Section of M. fragilis growing point图6 脆蔷薇珊瑚生长点纵切 |
Fig. 7 Montipora skeletal calcification process图7 蔷薇珊瑚骨骼钙化过程 |
Tab. 1 Weight and atomic percentage of each element in calcification process of skeleton表1 蔷薇珊瑚骨骼钙化过程各元素重量百分比和原子百分比 |
| 元素 | 重量比 | 原子比 | ||||||
|---|---|---|---|---|---|---|---|---|
| A | B | C | D | A | B | C | D | |
| C | 0 | 34.61 | 20.10 | 18.13 | 0 | 57.83 | 36.32 | 28.30 |
| O | 0 | 17.10 | 30.09 | 49.89 | 0 | 21.46 | 40.81 | 58.45 |
| Ca | 79.05 | 38.16 | 38.76 | 26.65 | 92.28 | 19.11 | 20.98 | 12.47 |
| I | 20.95 | 10.12 | 11.06 | 5.33 | 7.72 | 1.60 | 1.89 | 0.79 |
注: A为最初钙沉积点(生长点纵切); B为钙点扩大化(分枝中上部纵切); C为钙点平铺(分枝中下部纵切); D为形成羽簇(分枝底部纵切) |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}