
Journal of Tropical Oceanography >
Identification and phylogenetic analysis of Sagittidae (Chaetognatha) in the South China Sea based on mitochondrial CO#cod#x02160; gene
Received date: 2017-04-11
Request revised date: 2017-06-08
Online published: 2018-01-18
Supported by
Guangdong Natural Science Foundation (2014A030310495)
Director Fund of Marine Science and Technology, South China Sea Branch, State Oceanic Administration, People#cod#x02019;s Republic of China (1513)
Copyright
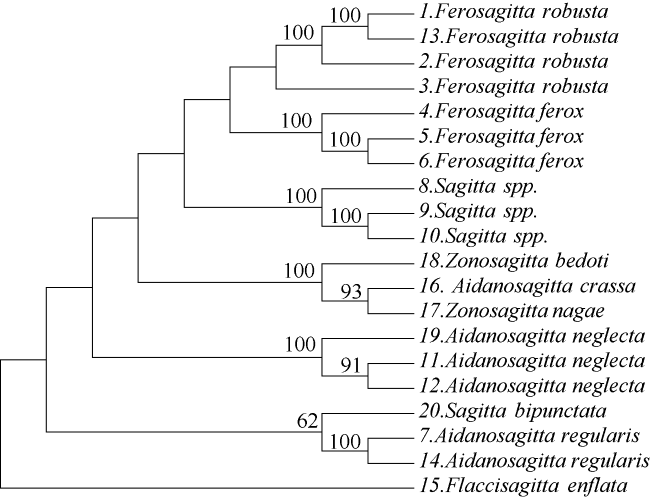
To improve the accuracy of Chaetognatha identification, we combined morphological identification and DNA barcoding based on the mitochondrial CO#cod#x02160;(cytochrome oxidase subunit #cod#x02160;) gene to identify some arrow worms from the northern South China Sea. The results showed that the morphologically and molecularly identified results were consistent, for the species of Ferosagitta robusta, Aidanosagitta neglecta and Aidanosagitta regularis. Furthermore, the neighbor-joining phylogenetic tree was constructed with the amplified CO#cod#x02160; sequences and those homologous sequences from GenBank based on Kimura 2-parameter, and the phylogenetic positions of Sagiitta (Sagittidae, Sagittoidea) were discussed. The results indicated that these CO#cod#x02160; sequences formed monophylotic clades compared with GenBank (91%~100%). The intraspecific genetic distance ranged from 0.000 to 0.015 with an average value of 0.004, while the interspecific genetic distance ranged from 0.081 to 0.173 with an average value of 0.127. The barcoding gap was detected between intra and interspecific variations, suggesting that the CO#cod#x02160; gene could be a standard DNA barcode for the Chaetognatha identification. This study was the first to apply DNA barcoding to phylogenetic analysis of Sagittidae, and to discuss the phylogenetic position at molecular level.
Key words: DNA barcoding; CO#cod#x02160;; gene; phylogeny; Chaetognatha; Sagittidae
CHEN Bin , HE Wei , LI Haitao , WU Peng , Xiao Yuzhang , LYU Xiangli , HE Jing . Identification and phylogenetic analysis of Sagittidae (Chaetognatha) in the South China Sea based on mitochondrial CO#cod#x02160; gene[J]. Journal of Tropical Oceanography, 2017 , 36(6) : 100 -108 . DOI: 10.11978/2017043
Tab. 1 Morphological characters of Sagittidae表1 箭虫的主要形态学特征 |
| 种名 | 数量 | 主要形态特征 |
|---|---|---|
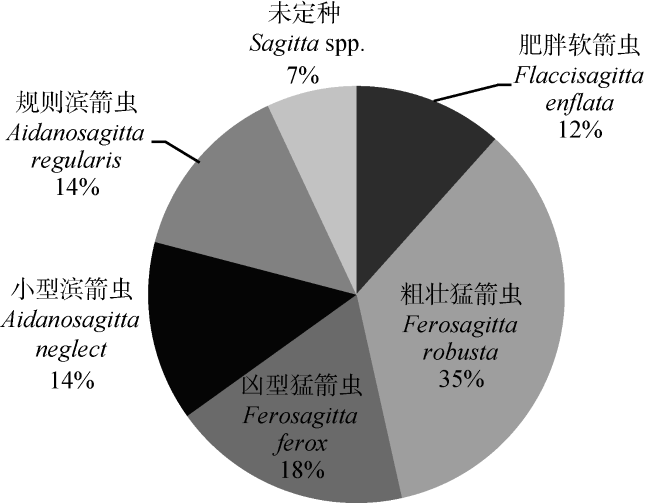
| 粗壮猛箭虫 Ferosagitta robusta | 15 | 体长10~12mm, 体坚硬, 不透明, 身体从头到尾隔的宽度相等。颈部明显, 领发达, 肠分歧, 卵巢宽大, 卵呈梨形, 储精囊呈倒梨型, 前鳍椭圆形, 后鳍呈圆形, 尾鳍呈三角形 |
| 肥胖软箭虫 Flaccisagitta enflata | 5 | 体长8~20mm, 体肥胖、柔软、透明, 颈部短, 无领, 肠不分歧, 卵巢短, 卵呈圆形, 储精囊呈球形, 前鳍仅边缘具鳍条, 尾鳍呈半圆形 |
| 凶型猛箭虫 Ferosagitta ferox | 8 | 体长9~16mm, 体坚实、不透明, 头大, 颈部明显, 肠分歧, 卵巢呈管状, 向前伸展可达颈部, 储精囊呈长卵圆形, 后鳍中央具一条狭小无鳍条带 |
| 规则滨箭虫 Aidanosagitta regularis | 6 | 体长6mm, 体细长, 较坚硬, 颈部不明显, 头到前鳍前半部的领非常发达, 其余为薄层延伸到尾, 卵巢前伸达腹神经节, 储精囊呈耳形 |
| 小型滨箭虫 Aidanosagitta neglecta | 6 | 体长6~9mm, 体坚硬、不透明, 颈部不明显, 领从颈到前鳍前端厚, 肠分歧, 卵巢充满体腔, 储精囊呈卵圆形, 接近后鳍但与尾鳍分离, 前后鳍均充满鳍条 |
| 未定种 Sagitta spp. | 3 | 体长15~16mm, 体坚实、不透明, 头大, 颈部明显。卵巢达前鳍中部, 卵大而圆, 储精囊呈耳状, 前鳍较后鳍长且大, 后鳍稍圆, 其中央无鳍条 |
Fig. 1 The proportion of Sagittidae图 1 箭虫科种类比例 |
Fig. 2 Features of Sagittidae.(a) Ferosagitta robusta, (b) Aidanosagitta neglecta, (c) Aidanosagitta regularis, (d) Ferosagitta ferox, (e) head of Ferosagitta ferox, (f) collarette of Aidanosagitta regularis, (g) seminal visical of Ferosagitta robusta, and (h) tale of Aidanosagitta regularis 图 2 部分箭虫外部形态 a. 粗壮猛箭虫; b. 小型滨箭虫; c. 规则滨箭虫; d. 凶型猛箭虫; e. 凶型猛箭虫头部; f. 规则滨箭虫泡状组织; g. 粗壮猛箭虫储精囊; h. 规则滨箭虫尾部 |
Tab. 2 Specimen information for which DNA barcodes were analyzed in this study, including phylogenetic position, species name and GenBank Accession Number表2 本研究涉及序列的名称、系统分类地位(科、属)及序列对应的GenBank登录号 |
| 序号 | 分类地位 | 中文种名 | 拉丁种名 | GenBank登录号 | 来源 |
|---|---|---|---|---|---|
| 1 | 箭虫科猛箭虫属 | 粗壮猛箭虫 | Ferosagitta robusta | KY882128 | 南海北部 |
| 2 | 箭虫科猛箭虫属 | 粗壮猛箭虫 | Ferosagitta robusta | KY882126 | 南海北部 |
| 3 | 箭虫科猛箭虫属 | 粗壮猛箭虫 | Ferosagitta robusta | KY882127 | 南海北部 |
| 4 | 箭虫科猛箭虫属 | 凶型猛箭虫 | Ferosagitta ferox | KY882123 | 南海北部 |
| 5 | 箭虫科猛箭虫属 | 凶型猛箭虫 | Ferosagitta ferox | KY882124 | 南海北部 |
| 6 | 箭虫科猛箭虫属 | 凶型猛箭虫 | Ferosagitta ferox | KY882125 | 南海北部 |
| 7 | 箭虫科猛箭虫属 | 规则滨箭虫 | Aidanosagitta regularis | KY882129 | 南海北部 |
| 8 | 未定 | 未定种 | Sagitta spp. | KY882132 | 南海北部 |
| 9 | 未定 | 未定种 | Sagitta spp. | KY882133 | 南海北部 |
| 10 | 未定 | 未定种 | Sagitta spp. | KY882134 | 南海北部 |
| 11 | 箭虫科滨箭虫属 | 小型滨箭虫 | Aidanosagitta neglecta | KY882130 | 南海北部 |
| 12 | 箭虫科滨箭虫属 | 小型滨箭虫 | Aidanosagitta neglecta | KY882131 | 南海北部 |
| 13 | 箭虫科猛箭虫属 | 粗壮猛箭虫 | Ferosagitta robusta | JN258035.1 | GenBank |
| 14 | 箭虫科滨箭虫属 | 正形滨箭虫 | Aidanosagitta regularis | FJ538304.1 | GenBank |
| 15 | 箭虫科软箭虫属 | 肥胖软箭虫 | Flaccisagitta enflata | KF977332 | GenBank |
| 16 | 箭虫科滨箭虫属 | 强壮滨箭虫 | Aidanosagitta crassa | HQ700947.1 | GenBank |
| 17 | 箭虫科带箭虫属 | 纳嘎带箭虫 | Zonosagitta nagae | NC013810.1 | GenBank |
| 18 | 箭虫科带箭虫属 | 百陶带箭虫 | Zonosagitta bedoti | JN258005.1 | GenBank |
| 19 | 箭虫科滨箭虫属 | 小型滨箭虫 | Aidanosagitta neglecta | JN258022 | GenBank |
| 20 | 箭虫科箭虫属 | 双斑箭虫 | Sagitta bipunctata | GQ368396 | GenBank |
Tab. 3 Distance estimation based on CO#cod#x02160; gene sequences using Kimura 2-parameter model表 3 基于CO#cod#x02160;基因序列通过K2P模型计算的箭虫科遗传距离 |
| 序号 | 种名 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ferosagitta robusta | ||||||||||||||||||||
| 2 | Ferosagitta robusta | 0.006 | |||||||||||||||||||
| 3 | Ferosagitta robusta | 0.002 | 0.004 | ||||||||||||||||||
| 4 | Ferosagitta robusta | 0.002 | 0.004 | 0.000 | |||||||||||||||||
| 5 | Ferosagitta ferox | 0.083 | 0.086 | 0.081 | 0.081 | ||||||||||||||||
| 6 | Ferosagitta ferox | 0.083 | 0.086 | 0.081 | 0.081 | 0.000 | |||||||||||||||
| 7 | Ferosagitta ferox | 0.083 | 0.086 | 0.081 | 0.081 | 0.000 | 0.000 | ||||||||||||||
| 8 | Aidanosagitta regularis | 0.141 | 0.143 | 0.143 | 0.143 | 0.160 | 0.160 | 0.160 | |||||||||||||
| 9 | Aidanosagitta regularis | 0.141 | 0.143 | 0.143 | 0.143 | 0.160 | 0.160 | 0.160 | 0.004 | ||||||||||||
| 10 | Aidanosagitta neglecta | 0.131 | 0.129 | 0.129 | 0.129 | 0.121 | 0.121 | 0.121 | 0.138 | 0.143 | |||||||||||
| 11 | Aidanosagitta neglecta | 0.131 | 0.129 | 0.129 | 0.129 | 0.121 | 0.121 | 0.121 | 0.138 | 0.143 | 0.000 | ||||||||||
| 12 | Aidanosagitta neglecta | 0.129 | 0.126 | 0.126 | 0.126 | 0.124 | 0.124 | 0.124 | 0.141 | 0.145 | 0.006 | 0.006 | |||||||||
| 13 | Aidanosagitta crassa | 0.131 | 0.129 | 0.133 | 0.133 | 0.099 | 0.099 | 0.099 | 0.138 | 0.143 | 0.124 | 0.124 | 0.126 | ||||||||
| 14 | Sagitta spp. | 0.092 | 0.090 | 0.094 | 0.094 | 0.105 | 0.105 | 0.105 | 0.141 | 0.141 | 0.136 | 0.136 | 0.138 | 0.103 | |||||||
| 15 | Sagitta spp. | 0.096 | 0.094 | 0.099 | 0.099 | 0.096 | 0.096 | 0.096 | 0.141 | 0.141 | 0.126 | 0.126 | 0.129 | 0.103 | 0.015 | ||||||
| 16 | Sagitta spp. | 0.092 | 0.090 | 0.094 | 0.094 | 0.105 | 0.105 | 0.105 | 0.141 | 0.141 | 0.136 | 0.136 | 0.138 | 0.103 | 0.000 | 0.015 | |||||
| 17 | Zonosagitta nagae | 0.136 | 0.143 | 0.138 | 0.138 | 0.112 | 0.112 | 0.112 | 0.124 | 0.129 | 0.119 | 0.119 | 0.126 | 0.092 | 0.121 | 0.121 | 0.121 | ||||
| 18 | Zonosagitta bedoti | 0.143 | 0.150 | 0.145 | 0.145 | 0.115 | 0.115 | 0.115 | 0.155 | 0.155 | 0.126 | 0.126 | 0.129 | 0.112 | 0.143 | 0.138 | 0.143 | 0.121 | |||
| 19 | Sagitta bipunctata | 0.129 | 0.126 | 0.126 | 0.126 | 0.143 | 0.143 | 0.143 | 0.121 | 0.126 | 0.141 | 0.141 | 0.138 | 0.126 | 0.148 | 0.143 | 0.148 | 0.145 | 0.143 | ||
| 20 | Flaccisagitta enflata | 0.143 | 0.150 | 0.145 | 0.145 | 0.173 | 0.173 | 0.173 | 0.170 | 0.170 | 0.150 | 0.150 | 0.153 | 0.160 | 0.148 | 0.153 | 0.148 | 0.160 | 0.168 | 0.153 |
Fig. 3 Phylogenetic tree based on CO#cod#x02160; gene sequences using Neighbor-joining method. No. 1~12 represent sequences obtained in this study, and No. 13~20 represent sequences obtained from GenBank图 3 基于CO#cod#x02160;基因序列通过邻接法构建的系统发育树(序号1~12为本研究测得的序列, 序号13~20为从Genbank获得的序列) |
| [1] |
DU FEIYAN, LI CHUNHOU, JIA XIAOPING, 2003. The situation and development trend of the study on Chaetognatha in China[J]. Journal of Shanghai Fisheries University, 12(1): 65-71 (in Chinese).
|
| [2] |
.
HUANG ZONGGUO, 2008. Marine species and their distribution in China[M]. Beijing: China Ocean Press: 698-699(in Chinese).
|
| [3] |
.
LIU RUIYU, 2008. Checklist of marine biota of China seas[M]. Beijing: Science Press: 842-843 (in Chinese).
|
| [4] |
.
XIAO YICHANG, 2004. Fauna sinica Vol.38[M]. Beijing: Science Press: 7-155 (in Chinese).
|
| [5] |
.中国科学院南海海洋研究所, 1983. 南海海洋生物研究论文集(一)[M]. 北京: 海洋出版社 17-63
South China Sea Institute of Oceanology, 1983. Contributions on marine biological research of the South China Sea[M]. Beijing: China Ocean Press: 17-63 (in Chinese).
|
| [6] |
.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
PEIJNENBURG K T C A,
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}