
Journal of Tropical Oceanography >
Genetic variations among Siganus oramin populations in coastal waters of southeast China based on mtDNA control region sequences
Received date: 2017-10-12
Request revised date: 2018-03-31
Online published: 2018-07-16
Supported by
Marine Fishery Science and Technology Industry Projects from Guangdong Province (A201601B10);Guangdong Provincial Key Laboratory of Fishery Ecology and Environment (FEEL-2017-13);Knowledge Innovation Basic Research Project of Shenzhen Science and Technology Program (JCYJ20170817103922921、JCYJ20170817103856495);South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Scientific Research Funds for Central Non-profit Institutes (2016TS01)
Copyright
In this study, we determined 828 bp mtDNA control region sequences of 100 Siganus oramin individuals collected in Daya Bay, Dapeng Bay, Shaba Bay, and Dongshan Bay where 72 variable sites defined 50 haplotypes. The parameter of haplotype diversity (h) were 0.893±0.053~0.957±0.030, and that of nucleotide diversity (π) was 0.0105±0.0031~0.0179± 0.0031, indicating a peculiarity of high haplotype diversity and low nucleotide diversity, The genetic diversity indexes of four geographical groups were, respectively, 0.0121 (DS), 0.0169 (DY), 0.0107 (DP), and 0.0184 (SB). A combination of the significant negative neutral test values of Fu’s Fs (-16.7079, p<0.01), the mismatch distribution pattern, and the network analysis revealed that a historical population expansion occurred at about 38300 years ago. The indices of genetic distance among populations (0.0112~0.0172) were at a similar level as the indices of genetic distance within populations (0.0107~0.0184). The overall genetic differentiation index (Fst) was 0.0066 (p>0.05), and the analysis of molecular variance showed that most of the genetic variations of the four populations was distributed within populations. With the above findings, it is suggested that S. oramin in the coastal waters of Southeast China is in low level of population genetic diversity and should receive effective protection.
Key words: mitochondrial D-loop; Siganus oramin; genetic diversity
HUANG Xiaolin , LI Wenjun , LIN Heizhao , YANG Yukai , LI Tao , YU Wei , HUANG Zhong . Genetic variations among Siganus oramin populations in coastal waters of southeast China based on mtDNA control region sequences[J]. Journal of Tropical Oceanography, 2018 , 37(4) : 45 -51 . DOI: 10.11978/2017109
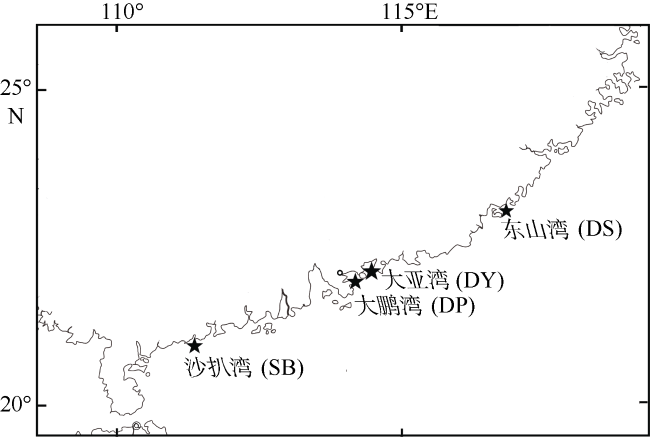
Fig. 1 Location of sampling of S. oramin图1 黄斑篮子鱼采集样点 |
Tab. 1 Genetic diversity parameters and neutral test in four populations of S. oramin表1 黄斑篮子鱼D-loop序列的不同地理群体的遗传多样性指数和中性检验 |
| 群体 | 样品数 | 单倍型 | 单倍型多样性 | 核苷酸多样性 | Tajima’s D | Fu’s Fs | ||
|---|---|---|---|---|---|---|---|---|
| D | p | Fs | p | |||||
| 大鹏湾(DP) | 25 | 15 | 0.893±0.053 | 0.0105±0.0031 | -1.2309 | 0.096 | -1.7442 | 0.249 |
| 东山湾(DS) | 25 | 16 | 0.850±0.073 | 0.0119±0.0033 | -0.6957 | 0.260 | -2.0316 | 0.213 |
| 沙扒湾(SB) | 25 | 19 | 0.957±0.030 | 0.0179±0.0031 | -0.2758 | 0.427 | -2.7962 | 0.156 |
| 大亚湾(DY) | 25 | 16 | 0.900±0.054 | 0.0164±0.0029 | 0.7290 | 0.800 | -0.6942 | 0.392 |
| 合计 | 100 | 50 | 0.900±0.027 | 0.0142±0.0016 | -0.5064 | 0.335 | -16.7079 | 0.001 |
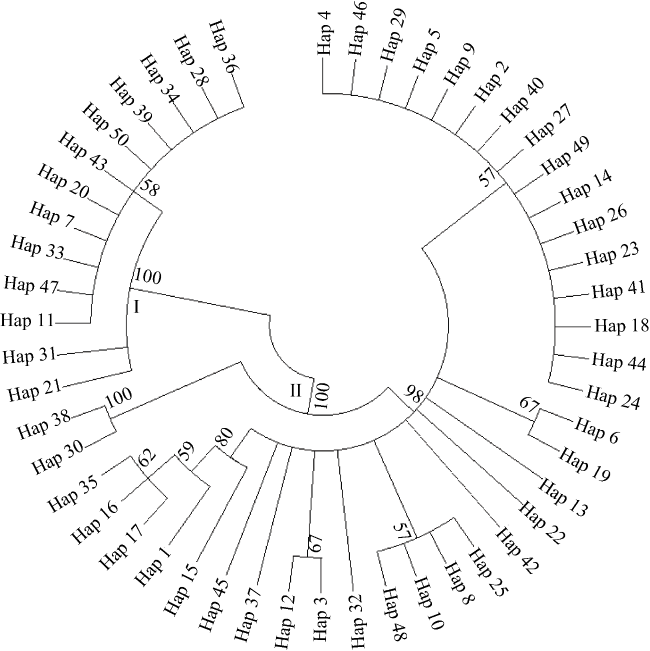
Fig. 2 Neighbor-joining tree constructed for mtDNA control region haplotypes of S. oramin图2 黄斑篮子鱼控制区单倍型的邻接系统树 |
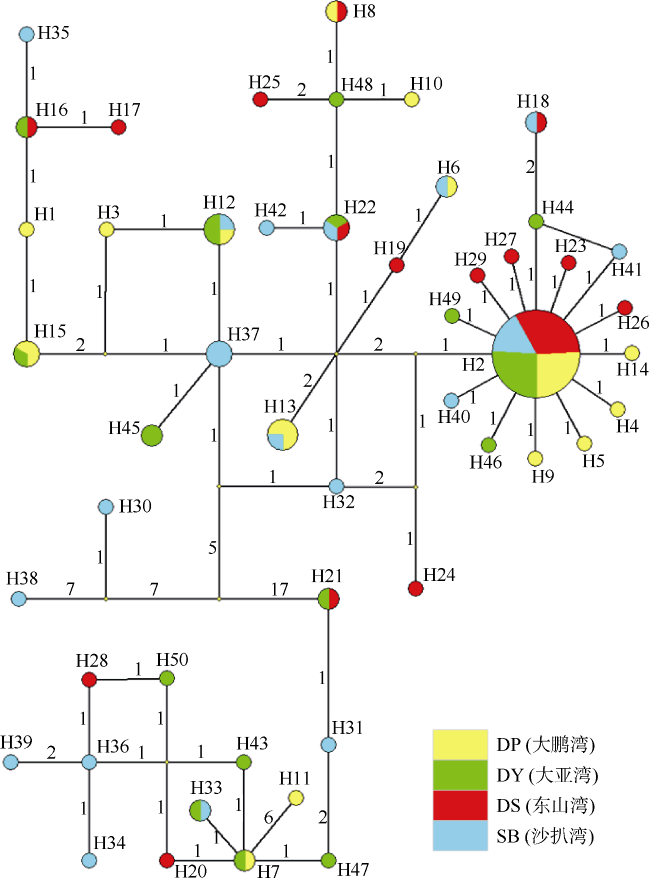
Fig. 3 The haplotype network of mtDNA control region sequences of four S. oramin populations图3 基于控制区序列构建的黄斑篮子鱼单倍型网络图 |
Tab. 2 Genetic distance between (low diagonal), within (diagonal) populations and Pairwis Fst表2 群体间遗传距离(对角线下方)、群体内遗传距离(对角线)和群体间(对角线上方)的Fst值 |
| 群体 | DP | DS | SB | DY |
|---|---|---|---|---|
| DP | 0.0107 | -0.0150 | 0.0320 | 0.0263 |
| DS | 0.0112 | 0.0121 | 0.0168 | 0.0045 |
| SB | 0.0150 | 0.0155 | 0.0184 | -0.0250 |
| DY | 0.0142 | 0.0146 | 0.0172 | 0.0169 |
注: 遗传距离p<0.01; p(Fst)>0.05; 加粗部分下方的遗传距离检验显著, 加粗部分和其上方的遗传分化指数不显著 |
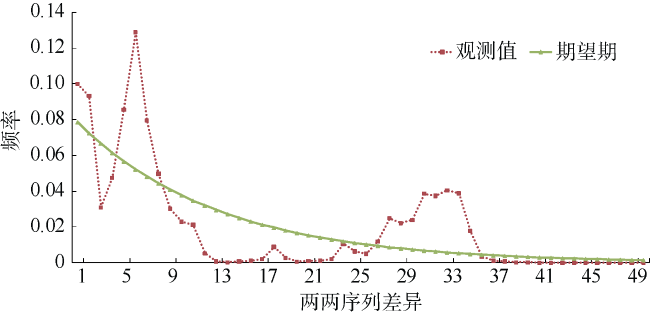
Tab. 3 Mismatch distribution parameters of S. oramin表3 核苷酸不配对分布分析参数估计值 |
| 群体 | 核苷酸不配对分布 | 拟合优度检验 | |||||
|---|---|---|---|---|---|---|---|
| 扩张 参数τ | 初始 值θ | 最终 值θ1 | 差平方 和SSD | p(SSD) | 粗糙指数R | p(R) | |
| DP | 6.5371 | 0 | 12.9541 | 0.0383 | 0.0500 | 0.0459 | 0.2400 |
| DS | 31.2402 | 0.0018 | 6.4972 | 0.0178 | 0.9500 | 0.0265 | 0.8100 |
| SB | 1.5000 | 13.0236 | 29.1961 | 0.0212 | 0.4700 | 0.0153 | 0.7200 |
| DY | 0 | 0 | 99999 | 0.8757 | 0 | 0.0304 | 1 |
| 合计 | 3.6582 | 6.0408 | 9.9870 | 0.0194 | 0.4100 | 0.0128 | 0.8400 |
Fig. 4 Mismatch-distribution analysis of S. oramin图4 核苷酸不配对分析图 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
中国科学院动物研究所, 中国科学院海洋研究所, 上海水产学院, 1962. 南海鱼类志[M]. 北京: 科学出版社: 733 (in Chinese).
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}