
Journal of Tropical Oceanography >
Study on the macrozoobenthic community structure in intertidal zone of Fangchenghe Estuary of Guangxi, China
Received date: 2018-05-31
Request revised date: 2018-08-29
Online published: 2019-01-16
Supported by
Science and Technology Project of State Oceanic Administration (YLFCJ20164006-F)
Science and Technology Project of Guangxi Oceanic Administration (GXHYJ100)
National Key Research and Development Program of China (2017YFC0506100)
Copyright
Based on the survey in May, August and November 2016 and February 2017, the effects of natural and anthropogenic factors on macrozoobenthic community structure in the intertidal zone of Fangchenghe Estuary of Guangxi, China were studied. A total of 252 species belonging to 10 phyla of intertidal macrozoobenthos were collected, among which Mollusca, Arthropoda and Annelida were richer in species abundance. Cerithidea cingulata, Ruditapes philippinarum, Mictyris longicarpus, Batillaria zonalis, Cyclina sinensis, Laternula nanhaiensis, Meretrix meretrix, and Gelolna coaxans were ranked as the top eight dominant species in this study area through the whole year. The annual average density and biomass of the study area were 203 ind•m-2 and 276.58 g•m-2, respectively, in which Mollusca occupied 69.58% and 83.73% to the total, respectively. ANOVA showed that salinity and sediment type factors had significant influences (p<0.05 and less) on macrozoobenthos density, biomass, species diversity H′, Richness d, Evenness J, and quadrat-based species abundance, while season and tidal zone factors had very small influences (p>0.05) on them. Salinity had larger influences on density and biomass than sediment type did, while sediment type had larger influences on species diversity H′, Richness d, Evenness J, and quadrat-based species abundance than salinity did. Both clustering analysis and multidimensional scaling analysis obtained the same results that the 21 sampling stations could be divided into three groups, basically owing to the effects from salinity and sediment type. The status of diversity index H′ showed that in general the intertidal macrozoobenthic communities were under middle-level fluctuation. Comprehensively, three factors, i.e., salinity, sediment type and anthropogenic fluctuation, played combined roles on the spatial distribution pattern of macrozoobenthic communities in the intertidal zone of Fangchenghe Estuary.
LAI Tinghe , HE Binyuan , HUANG Zhongjian , TANG Qiao , QIN Luyan , ZHU Ting , MO Zhenni , LIU Li , ZHONG Yunxu . Study on the macrozoobenthic community structure in intertidal zone of Fangchenghe Estuary of Guangxi, China[J]. Journal of Tropical Oceanography, 2019 , 38(2) : 67 -77 . DOI: 10.11978/2018034
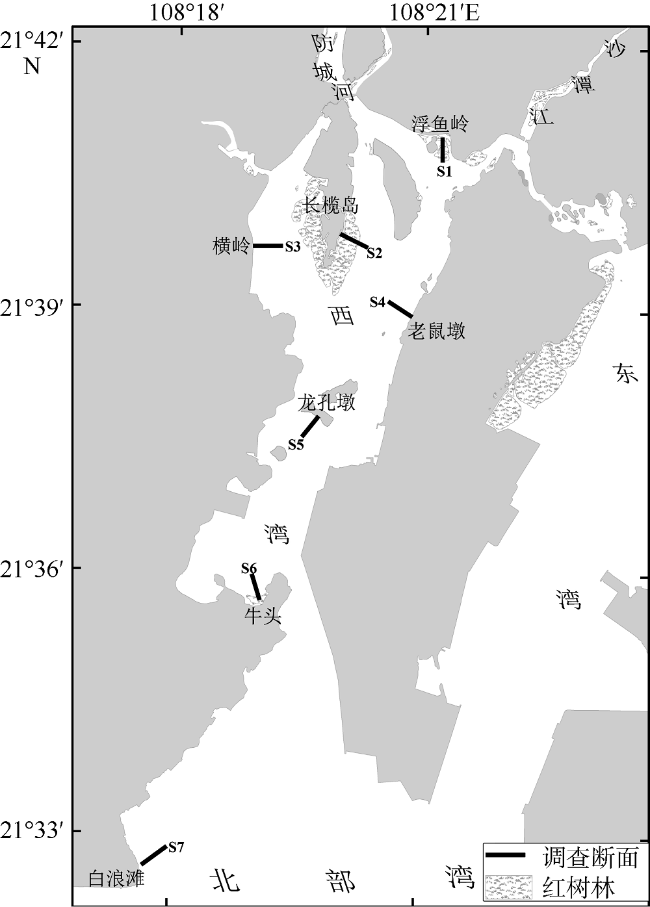
Fig. 1 Sampling stations for the macrozoobenthic community in the intertidal zone of Fangchenghe Estuary of Guangxi, China图1 防城河口湾潮间带大型底栖动物调查断面示意图 |
Tab. 1 Salinity, pH and habitat type at sampling stations for the intertidal macrozoobenthic community in Fangchenghe Estuary表1 防城河口湾潮间带大型底栖动物调查区域的盐度、pH和生境类型 |
| 断面 | 渗出水盐度/‰ | 上覆水盐度/‰ | 渗出水pH | 上覆水pH | 生境特点 |
|---|---|---|---|---|---|
| S1 | 17.6~27.2 | 3.9~27.0 | 7.1~7.4 | 7.2~8.1 | 高、中潮带为桐花树林, 低潮带为沙泥质裸滩 |
| S2 | 21.0~27.8 | 6.5~28.1 | 7.1~7.4 | 7.3~8.2 | 高潮带为秋茄林, 中潮带为白骨壤林, 低潮带为沙泥质裸滩 |
| S3 | 23.4~28.8 | 11.5~29.3 | 7.2~7.6 | 7.4~8.2 | 3个潮带均为淤泥质裸滩 |
| S4 | 22.0~28.3 | 10.6~29.0 | 7.2~7.6 | 7.5~8.2 | 高、中潮带为人工沙质裸滩, 低潮带为沙泥质裸滩 |
| S5 | 27.0~31.0 | 17.6~30.2 | 7.4~7.7 | 7.5~8.3 | 3个潮带均为沙泥质的滩涂贝类养殖场 |
| S6 | 27.4~32.0 | 19.3~30.8 | 7.3~7.8 | 7.8~8.5 | 高潮带为白骨壤林, 中、低潮带为沙泥质的滩涂贝类养殖场 |
| S7 | 28.0~33.0 | 27.0~31.8 | 7.6~7.9 | 7.8~8.6 | 3个潮带均为天然沙质裸滩 |
Tab. 2 Density and biomass of macrozoobenthic communities in the intertidal zone of Fangchenghe Estuary and the relative important values V (units: %) of dominant populations表2 防城河口湾潮间带大型底栖动物群落密度、生物量及优势种群相对重要值 |
| 断面 | 潮带 | 年均密度/(个•m-2) | 年均生物量/(g•m-2) | 优势种群及其相对优势度/% |
|---|---|---|---|---|
| S1 | 高潮带 | 244 | 252.59 | CEC 68.88, GEC 14.73, GLA 3.64, GLC 3.60, SER 2.08 |
| 中潮带 | 297 | 92.97 | MIL 70.09, CEC 15.42, SER 6.59, BAL 4.80 | |
| 低潮带 | 115 | 193.25 | PHS 57.5, CEC 15.2, LAT 5.5, LAN 4.4, CYS 3.7 | |
| 全断面 | 219 | 179.60 | CEC 54.86, MIL 17.15, GEC 7.72, SER 4.59, PHS 3.60 | |
| S2 | 高潮带 | 151 | 401.79 | CEC 61.29, GEC 32.30, CEM 3.86, ONV 1.17 |
| 中潮带 | 138 | 226.24 | CEC 66.30, MIL 11.95, LAT 11.24, UCV 4.53, GEC 4.21 | |
| 低潮带 | 264 | 295.18 | CEC 78.52, LAT 8.72, SER 3.01, UCV 2.16, BAL 2.09 | |
| 全断面 | 184 | 307.74 | CEC 77.95, GEC 10.29, LAT 5.14, MIL 1.68, UCV 1.49 | |
| S3 | 高潮带 | 68 | 201.29 | CYS 64.98, ANL 15.68, MSJ 8.30, CEC 8.22 |
| 中潮带 | 50 | 146.06 | ANL 43.16, TEG 24.02, PHS 13.57, TRS 9.09, PEV 6.01 | |
| 低潮带 | 47 | 119.54 | PHS 61.17, ANL 22.67, MSJ 8.54, PLD 2.40, MOP 1.26 | |
| 全断面 | 55 | 155.63 | ANL 39.14, PHS 22.77, CYS 19.73, MSJ 7.22, TEG 2.84 | |
| S4 | 高潮带 | 66 | 60.33 | MIL 85.66, DOW 12.35, MEM 1.74 |
| 中潮带 | 44 | 63.34 | MIL 90.34, DOW 3.88, CYS 2.94, LAT 2.17 | |
| 低潮带 | 138 | 108.82 | CEC 37.09, RUP 33.58, MEM 16.38, LAN 4.72, MIL 4.27 | |
| 全断面 | 83 | 77.50 | MIL 69.42, DOW 8.11, CEC 7.02, MEM 6.97, RUP 6.36 | |
| S5 | 高潮带 | 256 | 577.28 | CYS 65.01, CEC 24.26, BAZ 3.79, MEM 3.11, UCV 1.30 |
| 中潮带 | 229 | 380.61 | BAZ 37.11, CEC 34.30, BAC 9.95, CYS 7.41, SIN 7.29 | |
| 低潮带 | 560 | 1080.09 | RUP 91.78, LAN 7.04 | |
| 全断面 | 348 | 679.33 | RUP 75.80, CYS 5.98, LAN 5.90, CEC 4.96, BAZ 2.80 | |
| S6 | 高潮带 | 522 | 334.98 | CEC 75.82, BAZ 19.73, MIL 3.16 |
| 中潮带 | 325 | 380.64 | MOR 38.73, BAZ 23.90, CEC 15.17, MIL 12.23, MEM 6.36 | |
| 低潮带 | 242 | 277.66 | CEC 76.28, BAZ 21.56 | |
| 全断面 | 363 | 331.09 | CEC 66.17, BAZ 24.65, MIL 3.73, MOR 3.26 | |
| S7 | 高潮带 | 178 | 181.39 | CYC 60.96, MAV 24.72, MIL 8.27, MAA 5.86 |
| 中潮带 | 85 | 87.39 | CYC 41.93, MAA 41.29, CRP 8.81, DOW 5.91, MAV 1.05 | |
| 低潮带 | 252 | 346.68 | CRP 32.88, CHS 22.92, MEM 22.80, MOR 8.16, PAV 7.86 | |
| 全断面 | 172 | 205.15 | CYC 41.95, MAV 17.81, CRP 11.14, MAA 9.76, CHS 4.70 |
注: ANL=莱氏异额蟹Anomalifrons lightana, BAC=古氏滩栖螺Batillaria cumingi, BAL=潮间藤壶Balanus littoralis, BAZ=纵带滩栖螺Batillaria zonalis, CEC=珠带拟蟹守螺Cerithidea cingulata, CEM=小翼拟蟹守螺Cerithidea microptera, CHS =紫藤斧蛤Chion semigranosus, CRP=突畸心蛤Cryptonema producta, CYC=古明志圆蛤Cycladicama cumingii, CYS =青蛤 Cyclina sinensis, DIP=拟脊活额寄居蟹Diogenes paracristimanus, DOW=韦氏毛带蟹Dotilla wichmanni, FEP=长毛明对虾Fenneropenaeus penicillatus, GEC=红树蚬Gelolna coaxans, GLA=弓绿螂Glauconme cerea, GLA=白色吻沙蚕Glycera alba, GLC=中国绿螂Glauconme chinensis, ILN=宁波泥蟹Ilyoplax ningpoensis, LAN=南海鸭嘴蛤Laternula nanhaiensis, LAT=截形鸭嘴蛤Laternula truncata, LIA=亚氏海豆芽Lingula adamsi, MAA=高蛤蜊Mactra alta, MAV=四角蛤蜊Mactra veneriformis, MEL=长足长方蟹Metaplax longipes, MEM=文蛤Meretrix meretrix, MER=斧文蛤Meretrix lamarckii, MIL=长腕和尚蟹Mictyris longicarpus, MOP=菲律宾偏顶蛤Modiolus philippinarum, MOR=红明樱蛤Moerella rutila, MSJ=日本大眼蟹Macrophthalmus japonicus, NIJ=幼形亮樱蛤Nitidotellina juvenilis, NOL=背蚓虫Notomastus latericeus, ONV=石磺Onchididum verruculatus, PAV=南海毛满月蛤Pallucina vietnamica, PEV=翡翠贻贝Perna viridis, PHS=针骨沙鸡子Phyllophorus spiculatus, PLD=杜氏阔沙蚕Platynereis dumerilii, RUP=菲律宾蛤仔Ruditapes philippinarum, SER=斜肋齿蜷Sermyla riqueti, SIN=裸体方格星虫Sipunculus nudus, TEG=泥蚶Tegillarca granosa, TRS=亚光棱蛤Trapezium sublaevigatum, UCA=弧边招潮Uca arcuata, UCV=凹指招潮Uca vocans |
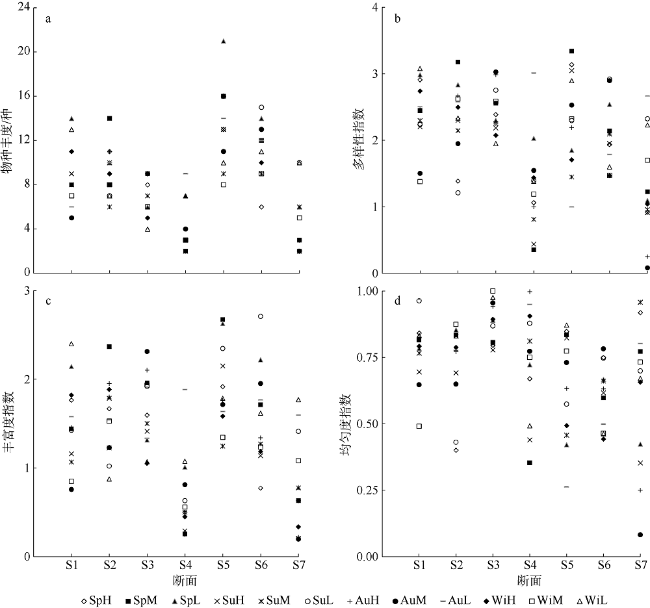
Fig. 2 Indexes of species abundance (a), species diversity H′ (b), Richness d (c), and Evenness J (d) of macrozoobenthic communities seasonally sampled in the intertidal zone of Fangchenghe Estuary. Among the three letters in each legend, the first two letters are the abbreviation of sampling season: Sp = Spring, Su = Summer, Au = Autumn, and Wi = Winter; and the last letter is the abbreviation of tidal zone: H = high tidal zone, M = middle tidal zone and L = low tidal zone图2 防城河口湾全年各潮带大型底栖动物群落物种丰度(a)、种类多样性指数(b)、丰富度指数(c)、均匀度(d) |
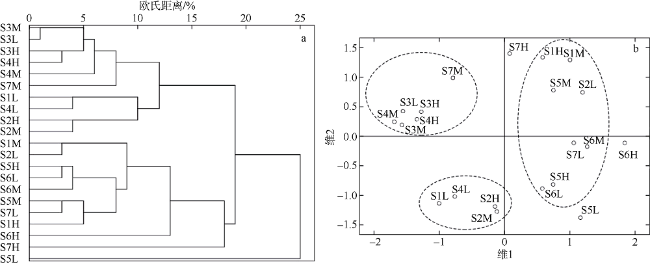
Fig. 3 The hierarchical cluster dendrogram (a) and 2-dimentional MDS ordinal configuration (b) of macrozoobenthic communities in the intertidal zone of Fangchenghe Estuary. H= high tidal zone, M= middle tidal zone and L= low tidal zone, Three dotted circles in figure b represent three major groups based on the MDS ordinal图3 防城河口湾潮间带大型底栖动物群落系统聚类(a)及MDS分析图(b) |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}