
Journal of Tropical Oceanography >
Dynamic characteristics of microtubule-dependent transport of Singapore Grouper Iridovirus in host cells
Received date: 2019-03-03
Request revised date: 2019-04-22
Online published: 2020-01-09
Supported by
National Natural Science Foundation of China(41606183)
Natural Science Foundation of Guangdong Province, China(2016A030310111)
National Key R&D Program of China(2017YFA0505300)
National Natural Science Foundation of China(21727816)
National Natural Science Foundation of China(21721003)
National Natural Science Foundation of China(21525314)
Copyright
Viruses are tiny intracellular parasites and rely on the cellular machinery for replication. Usually, internalized viruses hijack the microtubule for effectively moving from cell membrane to specific compartments for replication. However, the elaborate process of microtubule-dependent transport of viruses in live cells remains unclear. Singapore grouper iridovirus (SGIV) belongs to genus Ranavirus, family Iridoviridae, which is an important viral pathogens marine fish, and causes serious economic losses. In this study, different microtubule-dependent behaviors of SGIV were analyzed by the single-particle tracking technique. We found that some SGIV particles showed bi-directional and active motions from cell periphery to the microtubule organizing center (MTOC), with the highest speed reaching 0.2μm·s-1. When reaching the microtubule intersection, SGIV particles slowed down. Then, some virus was confined near the intersection, with an average speed of 0.008 μm·s-1. Other virus moved forward through the intersection, with a high speed of 0.2μm·s-1 again. In addition, SGIV infection could affect the morphology. The microtubule gradually formed a circular structure around the nucleus and viral factory during SGIV infection. These preliminary results reveal the complex interaction between SGIV and microtubule, further our understanding of intracellular activity of iridoviruses, and contribute to our understanding of the pathogenesis of iridoviruses infection.
WANG Liqun , WANG Shaowen , WANG Hongda , QIN Qiwei . Dynamic characteristics of microtubule-dependent transport of Singapore Grouper Iridovirus in host cells[J]. Journal of Tropical Oceanography, 2020 , 39(1) : 66 -73 . DOI: 10.11978/2019021
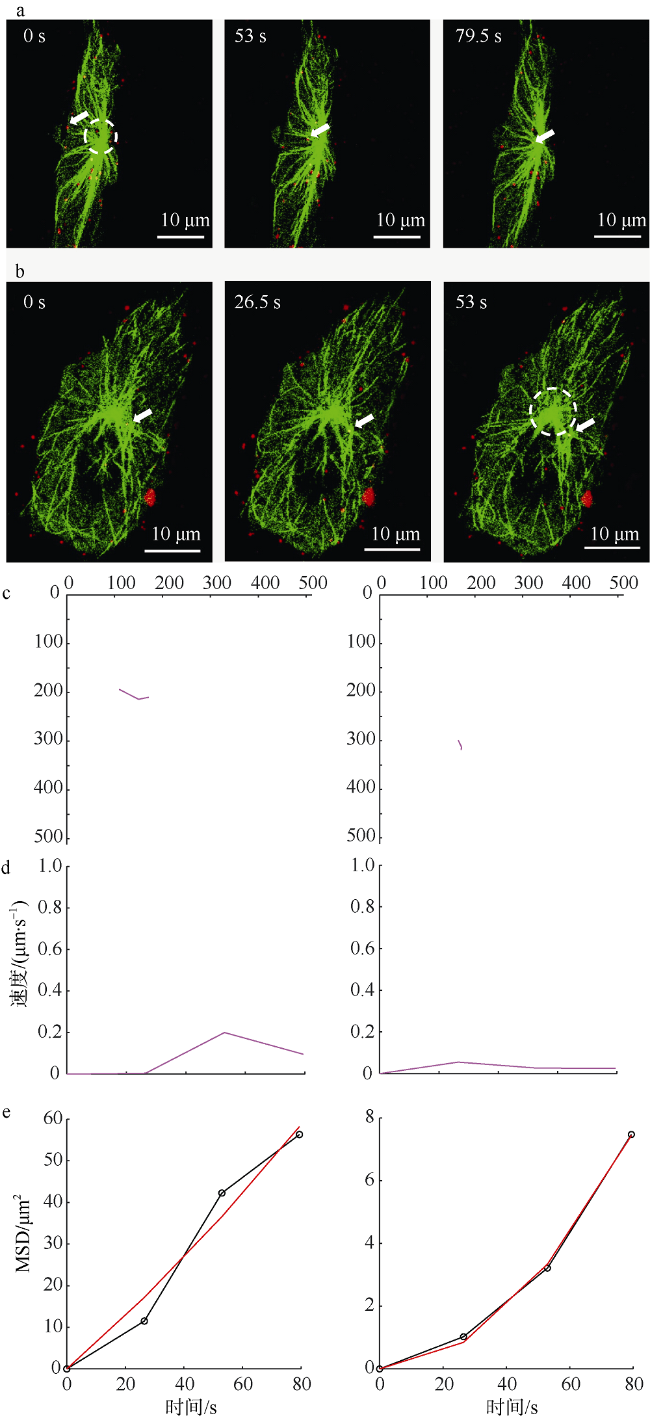
图1 SGIV沿微管的双向运动a. SGIV沿微管从细胞膜向MTOC运输的共聚焦图像; b. SGIV沿微管从MTOC向细胞膜运输的共聚焦图像。箭头指示正在沿微管运动的病毒粒子, 圆圈所示为MTOC。c. 图a和b所示病毒粒子的运动轨迹。d. 图a和b所示病毒粒子的瞬时运动速率。e. 图a和b图所示病毒粒子的MSD图; 图中红色曲线是公式LMSD=4Dt+(Vt)2的拟合, 其中LMSD是平均平方位移距离, D和V分别是扩散系数和拟合速度, t为时间; 黑色曲线是病毒粒子不同时间间隔的MSD数据 Fig. 1 Single virus moved bi-directionally along microtubule. a) Confocal images of single virus moved from cell membrane to MTOC along microtubules. Scale bar is 10 μm. The arrow indicates the virus particle moving on microtubule. The circle indicates MTOC. b) Confocal images of single virus moved from MTOC to cell membrane along microtubules. The arrow indicates the virus particle moving on microtubule. The circle indicates MTOC. c) The trajectory of the virus particle shown in (a) and (b). d) The instantaneous speed of the virus particle shown in (a) and (b). e) The MSD-time plots of the virus particle shown in (a) and (b). The red lines are fitted to the equation LMSD=4Dt+(Vt)2 (LMSD is the average square displacement distance, D and V are the diffusion efficient and mean speed of the particles, respectively). The black lines indicate the MSD data. |
图2 SGIV在微管交叉位置减速后继续运动a. SGIV在微管交叉位置减速后继续运动的共聚焦图像, 箭头指示沿微管运动的病毒粒子。b. 图a所示病毒粒子的运动轨迹。c. 图a所示病毒粒子的瞬时运动速率。d. 图a所示病毒粒子的MSD图 Fig. 2 Single virus decelerated near the intersection of microtubules and moved along the microtubule sequentially. a) Confocal images of single virus limited at an intersection of microtubules. The arrow indicates the virus particle moving on microtubule; b) the trajectory of the virus particle shown in (a); c) the instantaneous speed of the virus particle shown in (a); and d) the MSD-time plots of the virus particle shown in (a) |
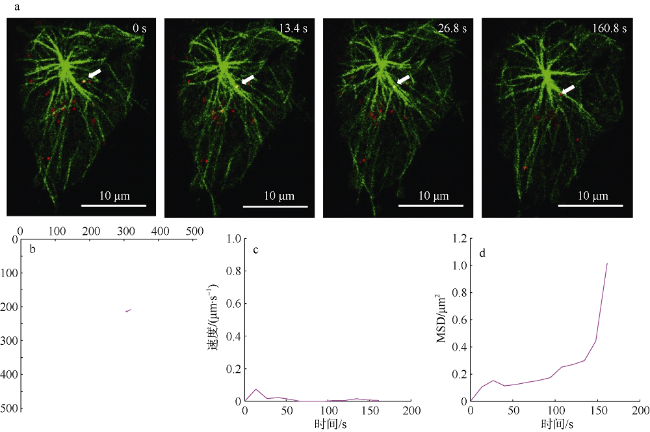
图3 SGIV在微管交叉处进行受限运动a. 单个SGIV病毒粒子在微管交叉处进行受限运动的共聚焦图像, 箭头指示沿微管运动的病毒粒子。b. 图a所示病毒粒子的运动轨迹。c. 图a所示病毒粒子的瞬时运动速率。d. 图a所示病毒粒子的MSD图 Fig. 3 Single virus limited at the intersection of microtubules. a) Confocal images of single virus limited at an intersection of microtubules. The arrow indicates the virus particle moving on microtubule; b) the trajectory of the virus particle shown in (a); c) the instantaneous speed of the virus particle shown in (a); and d) the MSD-time plots of the virus particle shown in (a) |
| [1] |
陈炜, 王嘉丽, 彭涛 , 等, 2005. 微管骨架在登革病毒感染ECV304细胞中作用的初步研究[J]. 第三军医大学学报, 27(7):628-631.
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}