
热带海洋学报 ›› 2025, Vol. 44 ›› Issue (1): 9-23.doi: 10.11978/2024043CSTR: 32234.14.2024043
印太交汇区八种鱼类群体遗传结构和连通性分析
黄红伟1,2,3( ), 张志新3,4, 仲嘉5, 林强3,4, 郭宝英1,2(), 严小军1,2()
), 张志新3,4, 仲嘉5, 林强3,4, 郭宝英1,2(), 严小军1,2()
- 1.浙江海洋大学, 海洋科学与技术学院, 浙江 舟山 316022
2.浙江海洋大学, 国家海洋设施养殖工程技术研究中心, 浙江 舟山 316022
3.中国科学院热带海洋生物资源与生态重点实验室, 中国科学院南海海洋研究所, 广东 广州 510301
4.中国科学院大学, 北京 100049
5.中国水产科学研究院长江水产研究所, 湖北 武汉 430223
-
收稿日期:2024-02-07修回日期:2024-03-13出版日期:2025-01-10发布日期:2025-02-10 -
通讯作者:郭宝英, 严小军 -
作者简介:黄红伟(1998—), 女, 硕士研究生, 从事海洋分子生态学研究。email: h18596254737@163.com
-
基金资助:国家重点研发计划项目(2022YFC3102403); 国家自然科学基金项目(42276126); 国家重点研发计划项目(2023YFC3108800)
The genetic structure and connectivity of eight fish species in the Indo-Pacific Convergence Region
HUANG Hongwei1,2,3(), ZHANG Zhixin3,4, ZHONG Jia5, LIN Qiang3,4, GUO Baoying1,2(), YAN Xiaojun1,2()
- 1. Marine Science and Technology College, Zhejiang Ocean University, Zhoushan 316022, China
2. National Engineering Research Center for Marine Aquaculture, Zhejiang Ocean University, Zhoushan 316022, China
3. Key Laboratory of Tropical Marine Bio-resources and Ecology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China
4. University of Chinese Academy of Sciences, Beijing 100049, China
5. Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430223, China
-
Received:2024-02-07Revised:2024-03-13Online:2025-01-10Published:2025-02-10 -
Contact:GUO Baoying, YAN Xiaojun -
Supported by:National Key Research and Development Program of China(2022YFC3102403); National Natural Science Foundation of China(42276126); National Key Research and Development Program of China(2023YFC3108800)
摘要:
印太交汇区是海洋生物多样性起源的中心, 具有极高的生物多样性。随着全球气候的持续变化和人类活动影响的日益加剧, 生态系统逐渐呈现破碎化, 此背景下, 海洋鱼类在连通破碎化的生态系统和维持生物多样性中发挥重要的生态功能。探究印太交汇区代表鱼类的遗传结构和连通性将有助于明确不同种群间的遗传多样性格局和扩散路径, 从而为生物多样性保护提供科学支撑。本研究在印太交汇区5个代表区域 (中沙群岛、南沙群岛、西沙群岛、海南岛和菲律宾) 收集了八种珊瑚礁栖鱼类 (日本刺尾鱼Acanthurus japonicus、波纹钩鳞鲀Balistapus undulatus、尾纹九棘鲈Cephalopholis urodeta、栉齿刺尾鲷Ctenochaetus striatus、金带齿颌鲷Gnathodentex aureolineatus、四带笛鲷Lutjanus kasmira, 、黑边角鳞鲀Melichthys vidua和多带副绯鲤Parupeneus multifasciatus) 共309个样本, 最终获得515条线粒体细胞色素C氧化酶亚基Ⅰ(cytochrome C oxidase subunit I, COI) 基因和 16S rDNA 序列; 为使研究覆盖范围更加广泛, 同时在 GenBank 下载了以上八种鱼类在中国台湾岛、南沙群岛、菲律宾、印度尼西亚海域的线粒体COI基因序列, 共86条。基于此, 研究分析了以上7个代表区域不同群体间的遗传多样性和遗传分化,结果显示八种鱼类在研究区域整体具有较高水平的遗传多样性和较低的遗传分化,其中多带副绯鲤和波纹钩鳞鲀最为明显; 印度尼西亚、中国台湾岛和南海诸岛群体同样具有较高的遗传多样性。研究进一步利用物种分布模型和最小成本路径模型探究了不同种群间的连通性, 在此基础上, 我们共收集了八种鱼类133047条分布信息, 使用最大熵算法(maximum entropy model, MaxEnt)预测当前气候下物种适宜分布的区域, 并利用ArcGis 10.2 软件 SDMtoolbox v2.5工具计算不同种群间的连通性。种群连通性揭示了菲律宾东岸—苏拉威西岛是以上八种鱼类在印太交汇区的重要扩散路径, 南海诸岛(南沙、中沙和西沙群岛)连接了中国台湾岛、菲律宾和印度尼西亚的不同群体, 起到了物种扩散的连通枢纽作用。综上所述, 菲律宾、印度尼西亚、中国台湾岛和南海诸岛应成为维持遗传多样性和保护连通性的优先保护区。本研究强调在生物多样性保护中应综合考虑种群连通性和遗传多样性的重要性, 通过结合宏观和微观层面的研究结果, 以期实现更为有效的生物多样性保护。
中图分类号:
- Q347
引用本文
黄红伟, 张志新, 仲嘉, 林强, 郭宝英, 严小军. 印太交汇区八种鱼类群体遗传结构和连通性分析[J]. 热带海洋学报, 2025, 44(1): 9-23.
HUANG Hongwei, ZHANG Zhixin, ZHONG Jia, LIN Qiang, GUO Baoying, YAN Xiaojun. The genetic structure and connectivity of eight fish species in the Indo-Pacific Convergence Region[J]. Journal of Tropical Oceanography, 2025, 44(1): 9-23.
表1
八种鱼类的样本数量和地理信息"
| 物种 | 自中沙群岛尾数 | 自西沙群岛尾数 | 自南沙群岛尾数 | 自海南岛尾数 | 自菲律宾尾数 |
|---|---|---|---|---|---|
| 日本刺尾鱼 | - | 7 | 10 | - | 6 |
| 波纹钩鳞鲀 | 9 | - | 12 | - | - |
| 尾纹九棘鲈 | 10 | 18 | 23 | - | - |
| 栉齿刺尾鲷 | - | 18 | 20 | - | - |
| 金带齿颌鲷 | 6 | 20 | 17 | - | - |
| 四带笛鲷 | - | 19 | 17 | - | - |
| 黑边角鳞鲀 | 25 | 10 | 22 | - | 10 |
| 多带副绯鲤 | 10 | 7 | 7 | 6 | - |
表1
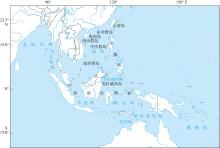
图1
研究区域地图 该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2021)5444号的标准地图制作, 底图无修改"

图1
表2
八种鱼类的地理分布数据(条)"
| 物种 | 下载数据 | 调查数据 | 文献检索数据 | 总数据 | 分析数据 |
|---|---|---|---|---|---|
| 日本刺尾鱼 | 170 | 25 | 4 | 199 | 12 |
| 波纹钩鳞鲀 | 15217 | 46 | 24 | 15287 | 158 |
| 尾纹九棘鲈 | 16650 | 51 | 17 | 16718 | 112 |
| 栉齿刺尾鲷 | 41773 | 52 | 10 | 41835 | 239 |
| 金带齿颌鲷 | 3492 | 62 | 1 | 3555 | 61 |
| 四带笛鲷 | 15315 | 0 | 0 | 15315 | 2027 |
| 黑边角鳞鲀 | 14562 | 62 | 1 | 14625 | 73 |
| 多带副绯鲤 | 25414 | 42 | 57 | 25513 | 238 |
表2
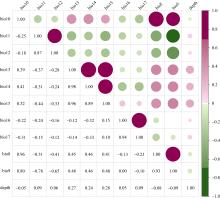
图2
11个海洋环境因子的共线性分析 bio8: 海表年平均盐度; bio9: 盐度最小月的海表年平均盐度; bio10: 盐度最大月的海表年平均盐度; bio11: 海表年平均盐度范围; bio12: 海表年平均盐度方差; bio13: 海表年平均温度; bio14: 温度最低月的海表年平均温度; bio15: 温度最高月的海表年平均温度; bio16: 海表年平均温度范围; bio17: 海表年平均温度方差; depth: 深度"

图2
表3
八种鱼类COI基因NCBI登录号"
| 物种 | 群体 | NCBI 登录号 | 物种 | 群体 | NCBI 登录号 |
|---|---|---|---|---|---|
| 日本刺尾鱼 | 菲律宾 | KY570696.1 | 黑边角鳞鲀 | 印度尼西亚 | GU673788.1 |
| 菲律宾 | FJ582663.1 | 印度尼西亚 | GU673787.1 | ||
| 中国台湾岛 | KU944984.1 | 印度尼西亚 | GU673786.1 | ||
| 波纹钩鳞鲀 | 印度尼西亚 | GU674269.1 | 印度尼西亚 | JN312832.1 | |
| 印度尼西亚 | GU674268.1 | 多带副绯鲤 | 印度尼西亚 | GU673922.1 | |
| 印度尼西亚 | GU673764.1 | 印度尼西亚 | GU673921.1 | ||
| 印度尼西亚 | GU673727.1 | 印度尼西亚 | MN870573.1 | ||
| 印度尼西亚 | GU673724.1 | 印度尼西亚 | MN870495.1 | ||
| 印度尼西亚 | JN312867.1 | 印度尼西亚 | MN870185.1 | ||
| 中国台湾岛 | KU945199.1 | 印度尼西亚 | MN870017.1 | ||
| 中国台湾岛 | KU945198.1 | 印度尼西亚 | MN869966.1 | ||
| 中国台湾岛 | KT718610.1 | 印度尼西亚 | MN869947.1 | ||
| 菲律宾 | FJ582893.1 | 印度尼西亚 | MN869941.1 | ||
| 尾纹九棘鲈 | 印度尼西亚 | MN870605.1 | 印度尼西亚 | JN312859.1 | |
| 印度尼西亚 | MN870316.1 | 南沙群岛 | KY371927.1 | ||
| 印度尼西亚 | MN870305.1 | 南沙群岛 | KY371926.1 | ||
| 印度尼西亚 | MN870241.1 | 南沙群岛 | KY371925.1 | ||
| 印度尼西亚 | MN869950.1 | 中国台湾岛 | KU944153.1 | ||
| 印度尼西亚 | JN313107.1 | 中国台湾岛 | KU944152.1 | ||
| 印度尼西亚 | JN312862.1 | 中国台湾岛 | KU944146.1 | ||
| 菲律宾 | KU064518.1 | 中国台湾岛 | KU944145.1 | ||
| 菲律宾 | KU064517.1 | 中国台湾岛 | KU944144.1 | ||
| 菲律宾 | KU064516.1 | 中国台湾岛 | KU944142.1 | ||
| 菲律宾 | KF009579.1 | 金带齿颌鲷 | 印度尼西亚 | GU674368.1 | |
| 菲律宾 | KF009578.1 | 印度尼西亚 | GU674365.1 | ||
| 菲律宾 | KC970463.1 | 印度尼西亚 | MN870418.1 | ||
| 菲律宾 | FJ583015.1 | 印度尼西亚 | MN869944.1 | ||
| 菲律宾 | FJ583014.1 | 南沙群岛 | KY371565.1 | ||
| 菲律宾 | FJ583013.1 | 南沙群岛 | KY371564.1 | ||
| 菲律宾 | FJ583012.1 | 南沙群岛 | KY371563.1 | ||
| 中国台湾岛 | KU943492.1 | 南沙群岛 | KY371562.1 | ||
| 中国台湾岛 | KU943491.1 | 南沙群岛 | KY371561.1 | ||
| 中国台湾岛 | KU943459.1 | 南沙群岛 | KY371560.1 | ||
| 中国台湾岛 | KU892843.1 | 南沙群岛 | KY371559.1 | ||
| 栉齿刺尾鲷 | 印度尼西亚 | GU673965.1 | 四带笛鲷 | 南沙群岛 | KY371696.1 |
| 印度尼西亚 | MN870174.1 | 南沙群岛 | KY371695.1 | ||
| 印度尼西亚 | MN870008.1 | 南沙群岛 | KY371694.1 | ||
| 印度尼西亚 | GU673967.1 | 南沙群岛 | KY371693.1 | ||
| 印度尼西亚 | JN311616.1 | 南沙群岛 | KY371692.1 | ||
| 中国台湾岛 | KU944980.1 | 南沙群岛 | KY371691.1 | ||
| 中国台湾岛 | KU893059.1 | 南沙群岛 | KY371690.1 | ||
| 中国台湾岛 | KU892976.1 | 南沙群岛 | KY371689.1 |
表3
表4
基于COI基因和16S序列的群体遗传多样性"
| 物种 | 群体 | 序列数(n) | 变异位点数目(S) | 单倍型数量(h) | 单倍型(基因) 多样性(Hd ) | 核苷酸多样性(π) | 核苷酸差异的平均数(K) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COI | 16S | COI | 16S | COI | 16S | COI | 16S | COI | 16S | COI | 16S | |||
| 日本 刺尾鱼 | 西沙群岛 | 7 | 6 | 2 | 1 | 2 | 2 | 0.571 | 0.333 | 0.002 | 0.001 | 1.143 | 0.333 | |
| 南沙群岛 | 7 | 10 | 6 | 2 | 4 | 3 | 0.810 | 0.378 | 0.004 | 0.001 | 2.381 | 0.400 | ||
| 菲律宾 | 8 | 4 | 6 | 112 | 6 | 4 | 0.893 | 1.000 | 0.004 | 0.135 | 2.071 | 74.000 | ||
| 整体 | 22 | 20 | 6 | 135 | 6 | 8 | 0.779 | 0.589 | 0.003 | 0.056 | 1.779 | 30.826 | ||
| 波纹 钩鳞鲀 | 中沙群岛 | 6 | 9 | 3 | 1 | 3 | 2 | 0.600 | 0.500 | 0.002 | 0.001 | 1.000 | 0.500 | |
| 南沙群岛 | 4 | 12 | 0 | 4 | 1 | 4 | 0.000 | 0.455 | 0.000 | 0.001 | 0.000 | 0.667 | ||
| 中国台湾岛 | 3 | - | 2 | - | 3 | - | 1.000 | - | 0.002 | - | 1.333 | - | ||
| 印度尼西亚 | 6 | - | 3 | - | 3 | - | 0.733 | - | 0.002 | - | 1.200 | - | ||
| 整体 | 19 | 21 | 113 | 5 | 8 | 5 | 0.795 | 0.486 | 0.105 | 0.001 | 57.924 | 0.638 | ||
| 尾纹 九棘鲈 | 中沙群岛 | 10 | 11 | 7 | 3 | 7 | 4 | 0.867 | 0.709 | 0.003 | 0.002 | 1.711 | 0.909 | |
| 西沙群岛 | 11 | 12 | 4 | 3 | 5 | 4 | 0.709 | 0.561 | 0.002 | 0.001 | 0.873 | 0.742 | ||
| 南沙群岛 | 19 | 23 | 50 | 88 | 11 | 7 | 0.830 | 0.806 | 0.030 | 0.024 | 15.930 | 13.024 | ||
| 菲律宾 | 10 | - | 5 | - | 5 | - | 0.756 | - | 0.003 | - | 1.489 | - | ||
| 中国台湾岛 | 4 | - | 5 | - | 2 | - | 0.500 | - | 0.005 | - | 2.500 | - | ||
| 印度尼西亚 | 7 | - | 5 | - | 6 | - | 0.952 | - | 0.004 | - | 2.095 | - | ||
| 整体 | 61 | 46 | 61 | 88 | 23 | 8 | 0.799 | 0.739 | 0.013 | 0.014 | 6.787 | 7.305 | ||
| 栉齿 刺尾鲷 | 西沙群岛 | 5 | 4 | 62 | 14 | 4 | 3 | 0.900 | 0.833 | 0.071 | 0.018 | 37.200 | 9.167 | |
| 南沙群岛 | 20 | 21 | 67 | 17 | 11 | 9 | 0.842 | 0.800 | 0.058 | 0.012 | 30.200 | 6.210 | ||
| 中国台湾岛 | 3 | - | 3 | - | 2 | - | 0.667 | - | 0.004 | - | 2.000 | - | ||
| 印度尼西亚 | 5 | - | 11 | - | 4 | - | 0.900 | - | 0.008 | - | 4.400 | - | ||
| 整体 | 33 | 25 | 77 | 18 | 17 | 10 | 0.822 | 0.787 | 0.050 | 0.013 | 26.426 | 6.500 | ||
| 金带 齿颌鲷 | 中沙群岛 | 6 | 6 | 1 | 0 | 2 | 1 | 0.333 | 0.000 | 0.001 | 0.000 | 0.333 | 0.000 | |
| 西沙群岛 | 5 | 20 | 0 | 1 | 1 | 2 | 0.000 | 0.100 | 0.000 | 0.000 | 0.000 | 0.100 | ||
| 南沙群岛 | 12 | 19 | 3 | 2 | 4 | 3 | 0.455 | 0.205 | 0.001 | 0.000 | 0.500 | 0.211 | ||
| 印度尼西亚 | 4 | - | 2 | - | 3 | - | 0.833 | - | 0.002 | - | 1.167 | - | ||
| 整体 | 27 | 45 | 4 | 3 | 5 | 4 | 0.393 | 0.130 | 0.001 | 0.000 | 0.484 | 0.133 | ||
| 四带笛鲷 | 西沙群岛 | 19 | 21 | 37 | 11 | 12 | 5 | 0.836 | 0.538 | 0.021 | 0.006 | 12.357 | 3.048 | |
| 南沙群岛 | 13 | 24 | 5 | 6 | 6 | 3 | 0.641 | 0.163 | 0.002 | 0.001 | 0.897 | 0.500 | ||
| 整体 | 32 | 45 | 41 | 17 | 16 | 7 | 0.756 | 0.358 | 0.014 | 0.003 | 8.482 | 1.859 | ||
| 黑边 角鳞鲀 | 中沙群岛 | 25 | 33 | 5 | 7 | 5 | 8 | 0.470 | 0.384 | 0.002 | 0.001 | 0.907 | 0.481 | |
| 西沙群岛 | 10 | 5 | 3 | 5 | 3 | 4 | 0.644 | 0.900 | 0.002 | 0.004 | 1.267 | 2.000 | ||
| 南沙群岛 | 15 | 27 | 6 | 11 | 4 | 9 | 0.600 | 0.604 | 0.003 | 0.002 | 1.429 | 1.083 | ||
| 菲律宾 | 11 | 10 | 6 | 5 | 5 | 5 | 0.618 | 0.667 | 0.002 | 0.002 | 1.091 | 1.000 | ||
| 印度尼西亚 | 4 | - | 3 | - | 3 | - | 0.833 | - | 0.003 | - | 1.500 | - | ||
| 整体 | 65 | 75 | 13 | 24 | 11 | 21 | 0.562 | 0.538 | 0.002 | 0.002 | 1.136 | 0.871 | ||
| 多带 副绯鲤 | 中沙群岛 | 10 | 10 | 3 | 17 | 4 | 10 | 0.533 | 1.000 | 0.002 | 0.012 | 0.867 | 6.911 | |
| 西沙群岛 | - | 3 | - | 9 | - | 3 | - | 1.000 | - | 0.010 | - | 6.000 | ||
| 南沙群岛 | 10 | 7 | 3 | 12 | 4 | 7 | 0.644 | 1.000 | 0.001 | 0.010 | 0.756 | 5.571 | ||
| 中国台湾岛 | 6 | - | 4 | - | 4 | - | 0.867 | - | 0.003 | - | 1.533 | - | ||
| 海南岛 | 6 | 6 | 4 | 15 | 3 | 6 | 0.600 | 1.000 | 0.002 | 0.011 | 1.333 | 6.533 | ||
| 印度尼西亚 | 10 | - | 7 | - | 7 | - | 0.911 | - | 0.004 | - | 1.978 | - | ||
| 整体 | 42 | 26 | 15 | 17 | 13 | 21 | 0.711 | 0.982 | 0.002 | 0.007 | 1.266 | 4.255 | ||
表4
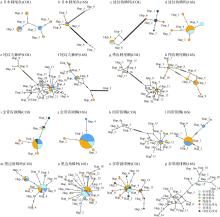
图3
基于COI基因和16S序列的TCS单倍型网络图"

图3

图4
基于COI基因和16S序列的K2P遗传距离分析 ZS: 中沙群岛; XS: 西沙群岛; NS: 南沙群岛; HN: 海南岛; TW: 中国台湾岛; PH: 菲律宾; Indonesia: 印度尼西亚。16S 序列: 对角线以上; COI 基因序列: 对角线以下"

图4
表5
基于Maxent模型评估8种鱼类的模型预测能力"
| 物种 | AUC (mean ± SD) | TSS (mean ± SD) | Boyce (mean ± SD) |
|---|---|---|---|
| 日本刺尾鱼 | 0.904 ± 0.093 | 0.806 ± 0.166 | 0.757 ± 0.263 |
| 波纹钩鳞鲀 | 0.988 ± 0.003 | 0.924 ± 0.030 | 0.882 ± 0.095 |
| 尾纹九棘鲈 | 0.986 ± 0.003 | 0.934 ± 0.017 | 0.847 ± 0.075 |
| 栉齿刺尾鲷 | 0.989 ± 0.005 | 0.934 ± 0.025 | 0.943 ± 0.046 |
| 金带齿颌鲷 | 0.983 ± 0.010 | 0.903 ± 0.053 | 0.811 ± 0.107 |
| 四带笛鲷 | 0.986 ± 0.003 | 0.907 ± 0.015 | 0.987 ± 0.008 |
| 黑边角鳞鲀 | 0.988 ± 0.007 | 0.912 ± 0.049 | 0.778 ± 0.100 |
| 多带副绯鲤 | 0.986 ± 0.007 | 0.944 ± 0.016 | 0.947 ± 0.032 |
表5

图5
当前气候条件下8种典型鱼类的适宜生境分布(颜色代表物种在印太交汇区的出现概率从强到弱) 该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663号的标准地图制作, 底图无修改"

图5
表6
基于Maxent模型预测8种鱼类的环境变量重要性"
| 物种 | 深度 | 海表年平均盐度 | 海表年平均盐度范围 | 海表年平均温度 | 海表年平均温度范围 |
|---|---|---|---|---|---|
| 日本刺尾鱼 | 1 | 0 | 0.428 | 0.026 | 0 |
| 波纹钩鳞鲀 | 1 | 0.138 | 0.065 | 0.025 | 0.233 |
| 尾纹九棘鲈 | 0.999 | 0.116 | 0.113 | 0.006 | 0.152 |
| 栉齿刺尾鲷 | 0.998 | 0.085 | 0.134 | 0.059 | 0.124 |
| 金带齿颌鲷 | 0.999 | 0.052 | 0.164 | 0.117 | 0.094 |
| 四带笛鲷 | 0.941 | 0.011 | 0.018 | 0.098 | 0.014 |
| 黑边角鳞鲀 | 0.988 | 0.100 | 0.172 | 0.031 | 0.380 |
| 多带副绯鲤 | 0.980 | 0.099 | 0.074 | 0.109 | 0.019 |
表6
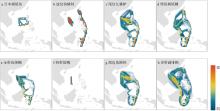
图6
基于 COI基因序列的种群连通性分析(颜色代表遗传连通性从强到弱) 该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663号的标准地图制作, 底图无修改"

图6
| [1] |
杜建国, 叶观琼, 周秋麟, 等, 2015. 近海海洋生态连通性研究进展[J]. 生态学报, 35(21): 6923-6933.
|
|
|
|
| [2] |
何思璇, 张典, 吴福星, 等, 2022. 厦门湾中华白海豚潜在生态廊道识别及人类活动干扰评估[J]. 生态学报, 42(21): 8555-8567.
|
|
|
|
| [3] |
李慧, 李丽, 吴巩胜, 等, 2018. 基于电路理论的滇金丝猴生境景观连通性分析[J]. 生态学报, 38(6): 2221-2228.
|
|
|
|
| [4] |
徐奎栋, 2021. 印太交汇区海洋生物多样性中心形成演化机制研究进展与展望[J]. 海洋与湖沼, 52(2): 262-273.
|
|
|
|
| [5] |
俞孔坚, 李迪华, 1997. 城乡与区域规划的景观生态模式[J]. 国外城市规划, (3): 27-31 (in Chinese).
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [1] | 黄雯, 冯逸, 李明, 武茜, 罗燕秋, 陈胤民, 王丽荣, 余克服. 造礁石珊瑚群体遗传学研究进展[J]. 热带海洋学报, 2024, 43(6): 13-26. |
| [2] | 钱钰坤, 刘统亚, 张华, 彭世球. 海洋中示踪物等值线的分形长度及其与混合效率的关系*[J]. 热带海洋学报, 2024, 43(1): 1-15. |
| [3] | 顾靖华, 朱建荣, 金智. 长江口宝钢码头溢油事故油膜漂移扩散数值模拟[J]. 热带海洋学报, 2022, 41(6): 159-170. |
| [4] | 李杨, 黄鹏起, 鲁远征, 屈玲, 郭双喜, 岑显荣, 周生启, 张佳政, 丘学林. 基于精细温度观测的南海东北部陆坡-深海盆底层湍流混合*[J]. 热带海洋学报, 2022, 41(1): 62-74. |
| [5] | 邹聪聪, 王丽娟, 吴志昊, 尤锋. 基于线粒体控制区序列的黄海日本鳀(Engraulis japonicus)的群体遗传结构*[J]. 热带海洋学报, 2021, 40(5): 25-35. |
| [6] | 李敏, 孔啸兰, 许友伟, 陈作志. 基于线粒体控制区序列的花斑蛇鲻遗传多态性分析[J]. 热带海洋学报, 2020, 39(4): 42-49. |
| [7] | 刘合香, 卢耀健, 王萌, 李广桃. 基于信息扩散技术的华南极端台风灾害风险评估[J]. 热带海洋学报, 2020, 39(3): 31-41. |
| [8] | 徐岩, 刘香全, 宋仁刚, 岑显荣, 郭双喜, 周生启. 扩散对流温盐台阶结构的数值模拟[J]. 热带海洋学报, 2019, 38(1): 11-18. |
| [9] | 黄小林, 李文俊, 林黑着, 杨育凯, 李涛, 虞为, 黄忠. 基于线粒体DNA D-loop序列的黄斑篮子鱼群体遗传多样性分析[J]. 热带海洋学报, 2018, 37(4): 45-51. |
| [10] | 赵志英, 梁丽运, 白丽蓉. 斑节对虾3个野生群体遗传多样性的微卫星标记分析[J]. 热带海洋学报, 2018, 37(3): 65-72. |
| [11] | 施晓峰, 苏永全, 王文成, 王军. 基于mtDNA控制区序列的3个黑棘鲷群体遗传结构特性研究[J]. 热带海洋学报, 2015, 34(1): 56-63. |
| [12] | 程香菊, 郭振仁, 刘国, 李兵. 大亚湾养殖区沉积物-水界面氮磷的释放通量及其在水体中的扩散迁移[J]. 热带海洋学报, 2014, 33(4): 77-84. |
| [13] | 邢元明, 杨磊,管玉平. 海表扩散层中气体行为的分子动力学模拟*[J]. 热带海洋学报, 2013, 32(2): 82-87. |
| [14] | 刘丽, 赵洁, 郭昱嵩, 刘楚吾. 4种笛鲷AFLP引物筛选及其遗传多样性分析[J]. 热带海洋学报, 2012, 31(4): 112-116. |
| [15] | 李莉好,喻达辉,黄桂菊,杜博,符云,童馨,郭奕惠,叶卫. 尼罗罗非鱼 Oreochromis niloticus 、 奥利亚罗非鱼 O. aureus 和红罗非鱼 O. sp. 群体遗传多样性的比较[J]. 热带海洋学报, 2012, 31(2): 102-109. |
|
||